
Метаболизм фруктозы и галактозы
Метаболизм фруктозы и галактозы включает пути использования их для синтеза других веществ (гетерополисахаридов, лактозы и др.) и участие в энергообеспечении организма. В последнем случае фруктоза и галактоза превращаются в печени либо в глюкозу, либо в промежуточные продукты её метаболизма. Таким образом, в результате фруктоза и галактоза наряду с глюкозой могут быть окислены до СО2 и Н2О или использованы на синтез гликогена и триацилглицеролов.
Причиной нарушения метаболизма фруктозы и галактозы может быть дефект ферментов, катализирующих промежуточные реакции их обмена. Эти нарушения встречаются относительно редко, но могут представлять достаточно серьёзную опасность, так как накапливаемые промежуточные метаболиты фруктозы и галактозы обладают токсичностью.
А. Метаболизм фруктозы
Значительное количество фруктозы, образующееся при расщеплении сахарозы, прежде чем поступить в систему воротной вены, превращается в глюкозу уже в клетках кишечника. Другая часть фруктозы всасывается с помощью белка-переносчика, т.е. путём облегчённой диффузии.
Метаболизм
фруктозы (рис. 7-69) начинается с реакции
фосфорилирования (реакция 1), катализируемой
фруктокиназой с образованием
фруктозо-1-фосфата. Фермент обнаружен
в печени, а также в почках и кишечнике.
Этот фермент обладает абсолютной
специфичностью, поэтому, в отличие от
глкжокиназы, инсулин не влияет на его
активность. Последнее обстоятельство
объясняет, почему уровень выведения
фруктозы в моче у больных сахарным
диабетом и здоровых не отличается.
Фруктозе-1-фосфат не может превращаться
во фруктозо-6-фосфат из-за отсутствия
соответствующего фермента. Вместо этого
фруктозо-1-фосфат далее расщепляется
фруктозо-1-фосфатальдолазой (альдолаза
В) на глицеральдегид и дигидроксиацетон-3-фосфат
(реакция 2).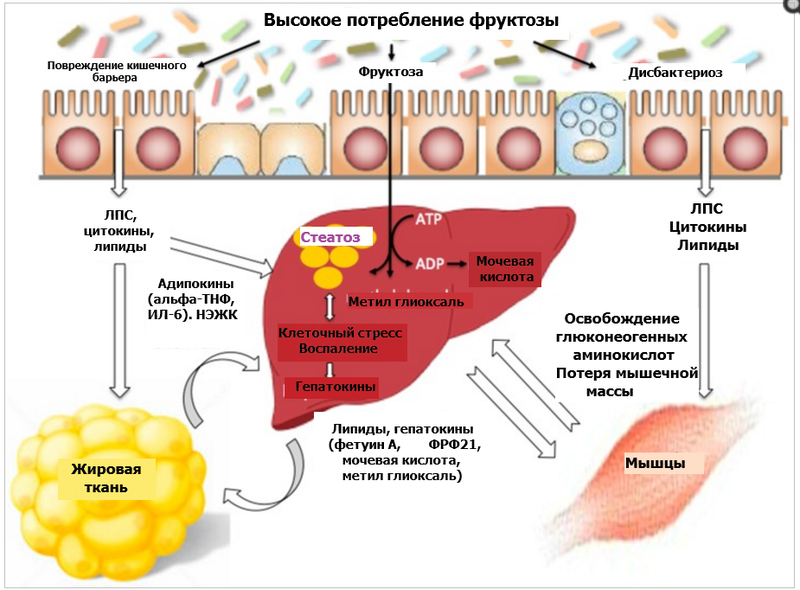
Следует отметить, что включение фруктозы в метаболизм через фруктозо-1-фосфат минует стадию, катализируемую фосфофруктокиназой (реакция 6), которая является пунктом метаболитического
364
Рис. 7-69. Метаболизм фруктозы. а — превращение фруктозы в дигидроксиацетон-3-фосфат и глицеральдегид-3-фосфат; б — путь включения фруктозы в гликолиз и глюконеогенез; в — путь включения фруктозы в синтез гликогена.
контроля скорости катаболизма глюкозы. Этим обстоятельством можно объяснить, почему увеличение количества фруктозы ускоряет в печени процессы, ведущие к синтезу жирных кислот, а также их этерификацию с образованием триацилглицеролов.
фруктоза метаболизм — Справочник химика 21
Эта первая стадия метаболизма состоит из 11 последовательных химических реакций, в которых глюкоза превращается во фруктозу, а затем в два производных глицеринового альдегида, содержащих три атома углерода. Лишь на одной-двух последних стадиях процесс разветвляется на различные маршруты, приводящие к пировиноградной кислоте, молочной кислоте, этанолу или ацетону. Каждая стадия гликолиза регулируется собственным катализатором, роль которого выполняет фермент с молекулярной массой 30000-500000.Далее фруктозо-1,6-бисфосфат может подвергаться дальнейшим превращением по пути гликолиза.
 Таков главный путь включения фруктозы в метаболизм мышечной ткани, почек, жировой ткани. [c.336]
Таков главный путь включения фруктозы в метаболизм мышечной ткани, почек, жировой ткани. [c.336] Развитие гипергликемии при диабете можно рассматривать также как результат возбуждения метаболических центров в ЦНС импульсами с хеморецепторов клеток, испытывающих энергетический голод в связи с недостаточным поступлением глюкозы в клетки ряда тканей. Роль системы фруктозо-2,6-бисфосфата в регуляции метаболизма углеводов, а также нарушения ее функционирования при сахарном диабете см. главу 16.
Известно также, что метаболизм фруктозы по гликолитическому пути в печени происходит гораздо быстрее, чем метаболизм глюкозы. Для метаболизма глюкозы характерна стадия, катализируемая фосфофруктокиназой-1. Как известно, на этой стадии осуществляется метаболический контроль скорости катаболизма глюкозы. Фруктоза минует эту стадию, что позволяет ей интенсифицировать в печени процессы метаболизма, ведущие к синтезу жирных кислот, их эстерификацию и секрецию липопротеинов очень низкой плотности в результате может увеличиваться концентрация триглицеридов в плазме крови.
Гидролиз фосфорорга-нических эфиров Расщепление фруктозо-дифосфата Zn метаболизм глюкозы Неизвестна [c.13]
Реакция, обратная реакции (3),—одна из основных реакций метаболизма. Вода в присутствии ионов (водн) расщепляет молекулу сахарозы на глюкозу и фруктозу. Этот процесс называется гидролизом, что означает .реакция с водой . [c.629]
Фруктоза трудно окисляемая, При ее восстановлении образуется смесь двух эпя-меров маннита и сорбита. Метаболизм фруктозы иллюстрирует схема, приведенная на рис. 59. [c.213]
Как уже отмечалось выше, сахароза потребляется не сама по себе, а сначала за пределами клеточной мембраны гидролизуется инвертазой до глюкозы и фруктозы. Инвертаза локализуется в клеточной стенке или в периплазматическом пространстве, в связи с чем сбраживание сахарозы в промышленном масштабе существенно отличается от сбраживания мальтозы.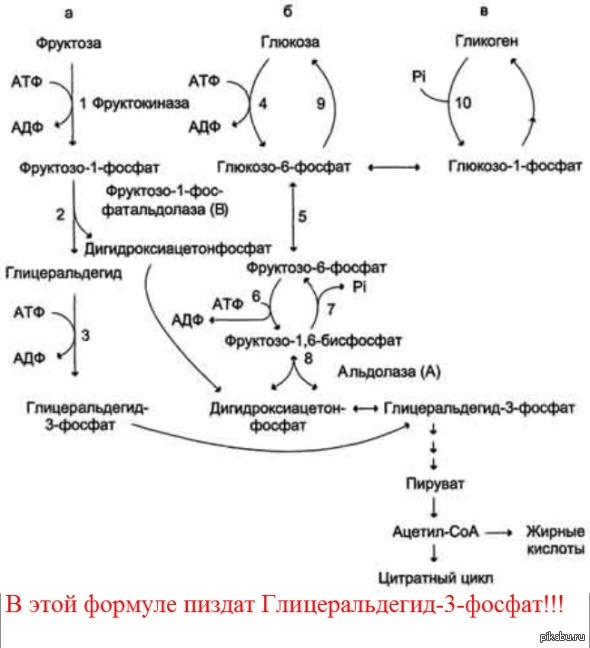 Глюкоза подавляет синтез инвертазы, а также метаболизм мальтозы на ранних стадиях брожения сусла. По мере уменьшения концентрации глюкозы в сусле подавление соответствующих генов прекращается и продуцируются ферментные системы для метаболизма этих дисахаридов (мальтозы и сахарозы). При повторном внесении таких дрожжей в свежее сусло с достаточной для подавления генов концентрацией глюкозы оба набора генов снова отключаются , внутри клетки активно расщепляются транспортер мальтозы и мальтаза, так что метаболизм мальтозы вскоре после внесения дрожжей прекращается. Инвертаза же, поскольку она локализована вне плазматической мембраны, не затрагивается этим регуляторным механизмом клеток, и способность их гидролизовать сахарозу сохраняется.
Глюкоза подавляет синтез инвертазы, а также метаболизм мальтозы на ранних стадиях брожения сусла. По мере уменьшения концентрации глюкозы в сусле подавление соответствующих генов прекращается и продуцируются ферментные системы для метаболизма этих дисахаридов (мальтозы и сахарозы). При повторном внесении таких дрожжей в свежее сусло с достаточной для подавления генов концентрацией глюкозы оба набора генов снова отключаются , внутри клетки активно расщепляются транспортер мальтозы и мальтаза, так что метаболизм мальтозы вскоре после внесения дрожжей прекращается. Инвертаза же, поскольку она локализована вне плазматической мембраны, не затрагивается этим регуляторным механизмом клеток, и способность их гидролизовать сахарозу сохраняется.
Фосфорилированные формы глюкозы и фруктозы в процессе их метаболизма способны взаимопревращаться, а также распадаться до триоз — фосфоглицеринового альдегида и фосфодиоксиацетона
При метаболизме фруктозы в организме большая ее часть фосфорилируется в печени при воздействии особого фермента — фруктокиназы в положении С-1 и образует фруктозо-1-фосфат. Считают, что при участии специфической альдолазы фруктозо-1-фосфат превращается в диокси-ацетонфосфат и глицеральдегид. Глицеральдегид восстанавливается до глицерина, затем через глицерол-З-фосфат окисляется до диоксиацетонфосфата. [c.126]
При образовании полисахаридов в клетках млекопитающих из фруктозы образуется фруктозо-6-фосфат, затем глюкозамин-6-фосфат и в конечном итоге — К -ацетилман-нозамин, иОР-Ы-ацетилглюкозамин, иОР-Ы-ацетилгалак-тозамин. Производные моносахаридов активно участвуют в метаболизме живой клетки, стимулируя процессы фотосинтеза, обеспечения клетки энергией, детоксикации и вывода ядовитых веществ, биосинтеза ароматических соединений, в том числе и аминокислот тирозина и фенилаланина, образования сложных биополимеров (полисахаридов, гликопротеинов, гликолипидов, нуклеиновых кислот).
Важная особенность П. ц. (в сравнении с др. путями метаболизма углеводов) — его гибкость. Если потребность в рибозо-5-фосфате значительно превышает потребность в НАДФН, то б.ч, глюкозо-б-фосфата по гликолитич. пути превращ. в глицеральдегид-З-фосфат, 1 молекула к-рого, аступля а р-ции е 2 молекулами фруктозо-6-фосфата,
Биосинтез Ф. тесно связан с метаболизмом сахарозы, к-рая служит как донором, так и акцептором остатков -D-фруктофуранозы. В клетках растений Ф. накапливаются в вакуолях и выполняют роль резервного материала (источник фруктозы), осморегулятора и антифриза. Содержание Ф. доходит до 30% от сухой массы в листьях, а в специализир. запасающих органах (корневищах, клубнях и луковицах) может превышать 60%. [c.191]
Глюконеогенез в печени сильно ускоряется глюкагоном и адреналином. Эффекты, вызываемые циклическим АМР, могут включать стимуляцию фруктозо-1,6-дифосфатазы и ингибирование фосфофруктокина-зы [46]. Влияние на взаимодействие между пируватом и РЕР, которое также имеет место, может быть непрямым и состоять в стимуляции а-кетоглутаратного метаболизма.
Вопрос о связи между действием фосфофруктокиназы и фруктозо-1,6-дифосфатазы [уравнение (11-19), стадия г рис. 11-11] остается нерешенным. Фруктозо-6-фосфат фосфорилируется и дает фруктозодифосфат, который в свою очередь гидролизуется, вновь превращаясь в фруктозо-6-фосфат. В результате получается бесполезный цикл (часто называемый бессмысленным циклом или субстратным циклом), который по существу ничем не завершается, кроме расщепления АТР до ADP и Р (АТРазная активность). Циклы этого типа часто встречаются в метаболизме, однако обычно они не приводят к гибельно быстрой потере АТР из-за четкого контроля метаболических процессов. В принципе в данный момент времени полностью активируются только один из двух ферментов, катализирующих стадию г [уравнение (11-19)]. В зависимости от метаболического состояния клетки может активно протекать процесс распада при небольшом биосинтезе или активный процесс биосинтеза при слабом распаде.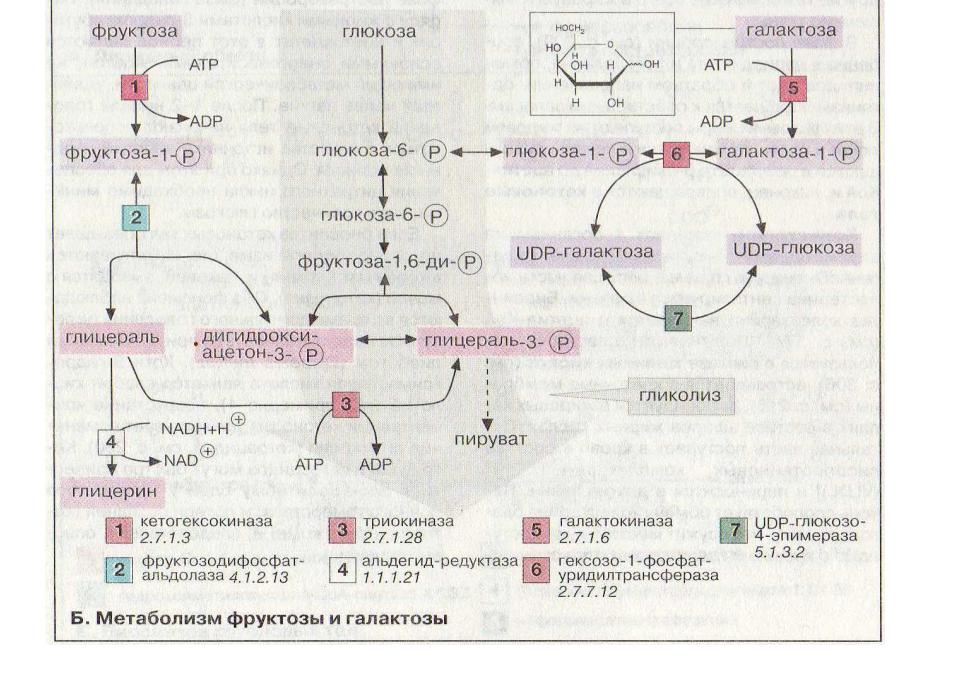 Некоторые из механизмов контроля показаны на рис. 11-11. Содержание АТР и АМР играет при этом наиболее важную роль—низкая концентрация АМР включает киназу и выключает фосфатазу. У разных видов ингибирующее действие по типу обратной связи может оказывать АТР, РЕР или цитрат. Не исключено, что в будущем будут обнаружены новые механизмы регуляции фруктозо-1,6-дифосфатазой. [c.513]
Некоторые из механизмов контроля показаны на рис. 11-11. Содержание АТР и АМР играет при этом наиболее важную роль—низкая концентрация АМР включает киназу и выключает фосфатазу. У разных видов ингибирующее действие по типу обратной связи может оказывать АТР, РЕР или цитрат. Не исключено, что в будущем будут обнаружены новые механизмы регуляции фруктозо-1,6-дифосфатазой. [c.513]
Значительная часть наших сведений о фотосинтезе была получена в результате изучения метаболизма двуокиси углерода, меченной С. Меченый углерод был обнаружен во многих продуктах, включая глюкозу, фруктозу и сахарозу. Для определения радиоактивности каждого из атомов углерода в данной молекуле была проведена деструкция ее на одиоуглеродиые фрагменты. [c.962]
Биосинтез декстрана идет следующим образом от молекулы сахарозы отщепляется молекула фруктозы, которая в своем метаболизме дает побочные продукты, такие, как молочную и уксусную кислоты или многоатомный спирт маннит. Оставшиеся молекулы глюкозы под действием фермента декстрансахаразы полимеризуются. Оптимум pH действия этого фермента от 5,2 до 5,6. Ферментация идет 20—24 ч (рис. 45). [c.137]
Далее трансальдолаза действует на продукты транскетолазной реакции, перенося Сз-фрагмент от молекулы С7-сахара на С3-М0-лекулу — 3-ФГА. В результате образуются молекулы Сй- и Сд-сахара. Один из продуктов реакции — фруктозо-6-фосфат является промежуточным соединением гликолитического пути, поэтому данная реакция есть вторая точка пересечения обоих путей углеводного метаболизма. Наконец, транскетолаза осуществляет перенос С2-фрагмента от молекулы Д-ксилулозо-5-фосфата на молекулу С4-сахара по той же схеме, что и в первой транскетолазной реакции. [c.257]
Известен ряд болезней, сопровождающихся отставанием скорости окисления пирувата от гликолиза. Так, при опухолевых заболеваниях гликолиз идет со скоростью, превышающей возможность цикла трикарбоновых кислот, что приводит к локальному повышению кислотности в опухолевой ткани эту особенность метаболизма рекомендуется использовать в терапии некоторых форм опухоли. Недостаточная активность ферментов, участвующих в метаболизме фруктозы и галактозы, приводит к таким метаболическим заболеваниям, как идеопатическая фруктозурия и галактоземия. [c.284]
Недостаточная активность ферментов, участвующих в метаболизме фруктозы и галактозы, приводит к таким метаболическим заболеваниям, как идеопатическая фруктозурия и галактоземия. [c.284]
При pH выше 5,0 D-арабино-З-гексулезо-б-фосфат может не пол-ностью изомеризоваться в фруктозо-6-фосфат и давать аллюлезо-6-фосфат, но это соединение далее в метаболизме этого цикла не участвует. [c.166]
Этот метод оказался весьма пригодным для объяснения биохимических проблем. Например, установлено, что метаболизм дезоксиинозина и дезоксиаденозина под действием свежих или консервированных эритроцитов вызывал накопление огромного количества дифосфата фруктозы, фосфата триозы и 5-фосфата дезоксирибозы. Кроме того, были обнаружены два неожиданных промежуточных соединения 1-фосфат 5-дезоксиксилулозы и 1,8-дифосфат 2-кето-5-дезоксиоктулозы [183]. Аналогичные примеры приведены в работах [184—187]. [c.122]
Измерение внутриклеточных концентраций метаболитов. Измерение концентраций промежуточных продуктов метаболизма в живой клетке сопряжено с большими экспериментальными трудностями. Поскольку клеточные ферменты катализируют быстро протекающие метаболические превращения, одна из обычных проблем при всяком экспериментальном вмешательстве в жизнь клетки связана с тем, что данные, полученные путем измерений, отражают не физиологические, а равновей1ые концентрации метаболитов. Поэтому любая экспериментальная методика будет надежной лишь в том случае, если с ее помощью удастся мгновенно подавить все ферментативные реакции в интактной ткани и тем самым предотвратить дальнейшие превращения промежуточных продуктов метаболизма. Этой цели можно достичь путем быстрого сжатия ткани между большими алюминиевыми пластинами, охлажденными жидким азотом ( —190°С) такой прием носит название фиксация замораживанием . После замораживания, мгновенно подавляющего действие ферментов, ткань растирают в порошок и ферменты инактивируют путем осаждения хлорной кислотой.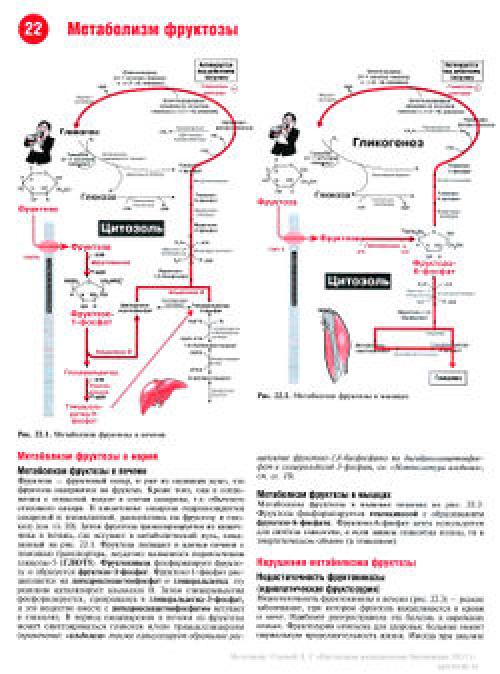 Осадок удаляют центрифугированием, а прозрачную надосадочную жидкость анализируют на содержание в ней метаболитов с помощью специфических ферментативных тестов. Истинную концентрацию данного метаболита в клетке определяют расчетным путем, учитывая общее содержание воды в ткани и данные измерений объема внеклеточного пространства, В табл. 1 приведены кажущиеся внутриклеточные концентрации субстратов и продуктов реакции фосфорилирования фруктозо-6-фосфата, катализируемой фер- [c.474]
Осадок удаляют центрифугированием, а прозрачную надосадочную жидкость анализируют на содержание в ней метаболитов с помощью специфических ферментативных тестов. Истинную концентрацию данного метаболита в клетке определяют расчетным путем, учитывая общее содержание воды в ткани и данные измерений объема внеклеточного пространства, В табл. 1 приведены кажущиеся внутриклеточные концентрации субстратов и продуктов реакции фосфорилирования фруктозо-6-фосфата, катализируемой фер- [c.474]
Большая часть потребленной свободной D-глюкозы в печени фосфорилируется при помощи АТР с образованием глюкозо-б-фосфата. Поглощенные в тонком кишечнике D-фруктоза, D-галактоза и D-манноза также превращаются в D-глюкозо-б-фосфат в результате ферментативного процесса, рассмотренного ранее (разд. 15.9). D-глюкозо-б-фосфат лежит, таким образом, на перекрестке всех путей превращения углеводов в печени. Метаболизм этого соединения в печени может осуществляться по пяти основным направлениям, и выбор какого-нибудь одного из них зависит от ежечасно и даже ежеминутно меняющихся спроса и предложения (рис. 24-9). [c.752]
Цикл начинается с того, что под действием АТФ, образовавшегося в фотохимических реакциях, рибулозо-5-фосфат превращается в дифосфат. Последний соединяется с диоксидом углерода с образованием неустойчивого шестиуглеродного соединения, которое дает две молекулы глицериновой кислоты-З-фос-фата. Для ее восстановления в глицеральдегид-З-фосфат и изомерный ему диоксиацетонфосфат необходимы тйкже АТФ и НАДФН. Два последних триуглеродных соединения превращаются во фруктозо-6-фосфат, который далее претерпевает обычный метаболизм углеводов (гл. 15). Кроме этого, углеводы могут запасаться в виде сахарозы или крахмала. [c.290]
Главный альтернативный путь метаболизма глюкозы и фруктозы известен под названием пентозофосфатный цикл или гексозомонофосфатный шунт. Фруктозо-6-фосфат изомеризуется в глюкозо-6-фосфат, который окисляется до глюконовой кис-лоты-6-фосфата (Се).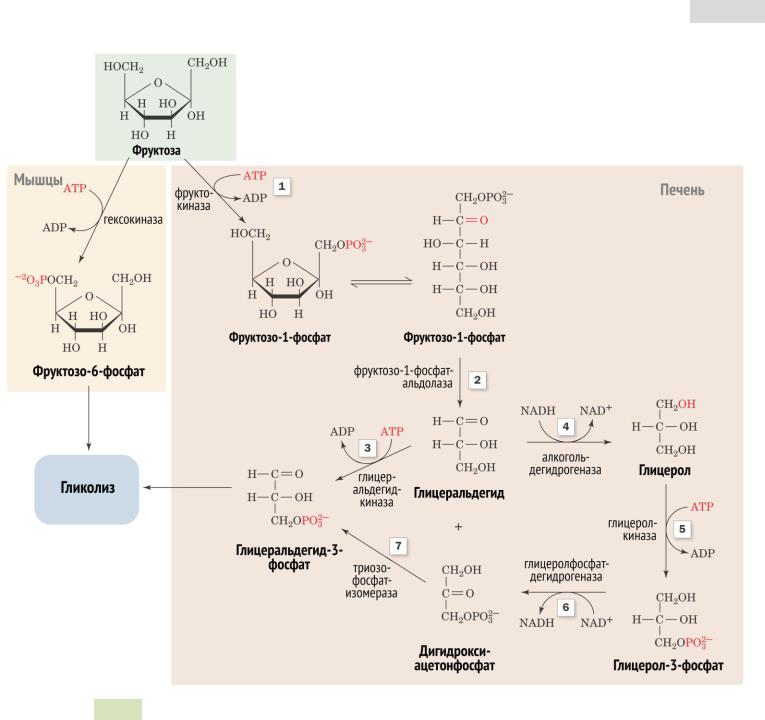 Это соединение теряет диоксид углерода, переходя в рибулозо-5-фосфат (С5). Во время этих процессов окисления образуется 2 моля НАДФН (рис. 15.6), который может включаться в цикл дыхания (разд. 13.3). [c.313]
Это соединение теряет диоксид углерода, переходя в рибулозо-5-фосфат (С5). Во время этих процессов окисления образуется 2 моля НАДФН (рис. 15.6), который может включаться в цикл дыхания (разд. 13.3). [c.313]
Метаболиты. Уже при поверхностном знакомстве с веществами, участвующими в клеточном метаболизме, легко заметить, что многие из них находятся в фосфорилированном состоянии, т.е. в форме эфиров фосфорной кислоты, Нефосфорилированные промежуточные продукты содержат карбоксильные группы или ионизующиеся основные группы. Создается впечатление, что ферменты могут воздействовать только на те метаболиты, в которых имеется ионизированная, т.е. заряженная, группа. Незаряженные молекулы или группы всегда связаны с кофер-ментами или простетическими группами ферментов некоторые образуют шиффовы основания с диаминокислотой лизином, содержащейся в активном центре ферментного белка. Неионизррованными бывают лишь соединения, стоящие в начале и в конце тех или иных метаболических путей таковы многие субстраты и некоторые выделяемые клеткой продукты обмена (глюкоза, фруктоза, этанол, ацетон, 2-пропанол, бутанол, глицерол и т.д.). Остается пока открытым вопрос о том, связано ли наличие ионизированных промежуточных соединений с функциями ферментов или с особой способностью клетки удерживать такие метаболиты. [c.220]
Как было показано ранее, фосфоглицериновая кислота восстанавливается до фосфоглицеральдегида под действием АТФ и восстановленного никотинамида. Образовавшийся глицеральдегид может продолжать поддерживать процесс анаэробного метаболизма, идущего через образование фруктозы и глюкозы и заканчивающегося образованием запаса пищи в виде крахмала или в виде целлюлозы, откладывающейся в стенке клетки как крахмал, так и целлюлоза являются полисахаридами. [c.41]
Различные гексозамины, входящие в состав многих мукополисахаридов и глюкопротеидов, а также полисахаридов оболочки бактериальных клеток, в процессе метаболизма берут свое начало от D-фруктозо-б-фосфата. Это соединение превращается в a-D-глюкозамин при взаимодействии с амидной КНо-группой глутамина, играющей роль источника азота [c.308]
Это соединение превращается в a-D-глюкозамин при взаимодействии с амидной КНо-группой глутамина, играющей роль источника азота [c.308]
Д-Глицеральдегид-З-фосфат (79) является важнейшим веществом в промежуточном метаболизме. Его ( )-модификацию можно получить из диэтнлацеталя глицидного альдегида (80) реакцией с дизал1ещениым фосфатом калия и последующим удалением ацетальной группы водной кислотой [256]. Д-Глицеральдегид-З-фос-фат образуется прп фотосинтезе [257] на очень ранней стадии фиксации углерода путем восстаиовлеиия 1,3-фосфата глицериновой кислоты. Он образуется также при гликолизе за счет рет-роальдольного расщепления 1,6-дифосфата фруктозы с участием фермента альдолазы, которое приводит к смеси О-глицеральдегид- [c.558]
Так, при действии на сахарозу фермента пекарских дрожжей были получены два различных олигосахарида —тураноза (I) [О-а-О-глюко-пиранозил-(1 3)-Б-фруктофураноза] и мальтулоза (II) [О-а-О-глюко-пиранозил-(1- 4)-0-фруктоза]. При изучении метаболизма сахарозы [c.38]
Обьгано около 90% всех переносимых по флоэме питательных веществ составляет дисахарид глюкоза. Это сравнительно инертный и хорошо растворимый углевод, который не играет почти никакой роли в метаболизме и поэтому служит идеальной транспортной формой, так как маловероятно, чтобы он расходовался в процессе переноса. Основное предназначение сахарозы — вновь превратиться в более активные моносахариды — глюкозу и фруктозу. Высокая растворимость позволяет ей достигать во фло-эмном соке очень высокой концентрации, например у сахарного тростника она составляет до 25% (масса/объем). [c.129]
Со стороны растения-хозяина энергетика клубеньков обеспечивается синтезом клубенек-специфичных изоформ ферментов С-метаболизма (табл. 4.5, рис. 4.5). Среди них одно из центральных мест занимает саха-розосинтаза (88), гидролизующая сахарозу с участием уридиндифосфата. В клубеньках сои 88 составляет 3—4 % от общего количества растворимых белков цитозоля. Образующиеся в результате гидролиза гексозы (глюкоза и фруктоза) претерпевают обычный путь катаболизма — гликолиз. Альтернативный путь — пентозофосфатный цикл, по-видимому, имеет в клубеньках второстепенное значение. [c.180]
Образующиеся в результате гидролиза гексозы (глюкоза и фруктоза) претерпевают обычный путь катаболизма — гликолиз. Альтернативный путь — пентозофосфатный цикл, по-видимому, имеет в клубеньках второстепенное значение. [c.180]
— взаимопревращение — Биохимия
Поскольку в кишечнике всасываются все поступающие с пищей моносахариды (фруктоза, галактоза, манноза и т.п.), то перед организмом встает задача превратить полученные гексозы в глюкозу для ее дальнейшего использования в реакциях метаболизма – происходит превращение сахаров. При дефекте соответствующих ферментов возникает накопление моносахаридов в крови – галактоземия и фруктоземия.
Превращение моносахаров
Цель этого процеса – создание только одного субстрата для реакций метаболизма, а именно α-D-глюкозы, что позволяет сэкономить ресурсы, не образовывать множество ферментов для каждого вида моносахарида. Реакции образования свободной глюкозы протекают в эпителии кишечника и, в основном, в гепатоцитах.
У детей некоторое время после рождения, даже при гипогликемии, в крови отмечается относительный избыток других моносахаридов, например, фруктозы и галактозы, что обычно связано с функциональной незрелостью печени.
Превращение галактозы
Галактоза сначала подвергается фосфорилированию по 1-му атому углерода. Отличительной особенностью является превращение в глюкозу не напрямую, а через синтез УДФ-галактозы из галактозо-1-фосфата. Источником УМФ является УДФ-глюкоза, имеющаяся в клетке. Образованная УДФ-галактоза впоследствии изомеризуется в УДФ-глюкозу и далее ее судьба различна.
Она может:
- участвовать в реакции переноса УМФ на галактозо-1-фосфат,
- превращаться в свободную глюкозу и выходить в кровь,
- отправляться на синтез гликогена.
Превращение галактозы в глюкозу
(обратимость обеих уридил-трансферазных реакций не показана)
Биохимическое усложнение вроде бы простой реакции эпимеризации вызвано, видимо, синтезом УДФ-галактозы из глюкозы в молочной железе для получения лактозы при образовании молока.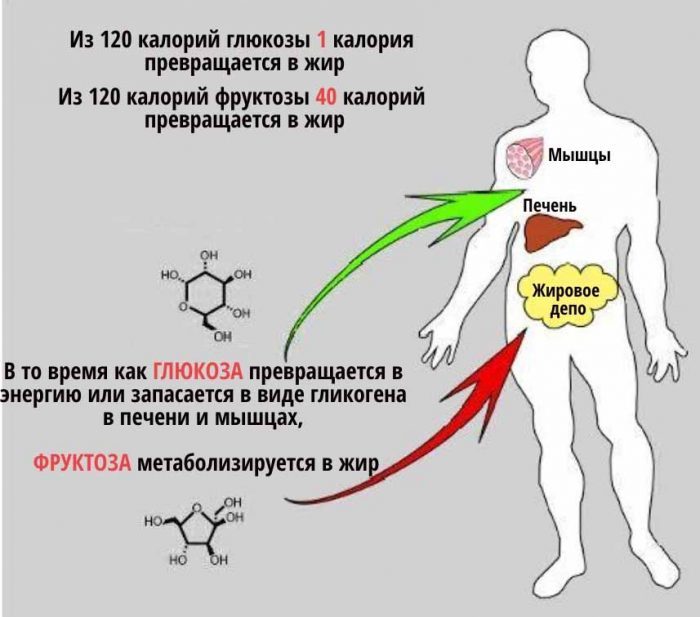 Также галактоза используется при синтезе соответствующих гексозаминов в гетерополисахаридах.
Также галактоза используется при синтезе соответствующих гексозаминов в гетерополисахаридах.
Нарушения превращения галактозы
Нарушения обмена галактозы могут быть вызваны генетическим дефектом одного из ферментов:
- галактокиназы, частота дефекта 1:500000,
- галактозо-1-фосфат-уридилтрансферазы, частота дефекта 1:40000,
- эпимеразы, частота дефекта менее 1:1000000.
Заболевание, возникающее при этих нарушениях, получило название галактоземия.
Диагностика. Дети отказываются от еды. Концентрация галактозы в крови возрастает до 11,1-16,6 ммоль/л (норма 0,3-0,5 ммоль/л), в крови появляется галактозо-1-фосфат. К лабораторным критериям относятся также билирубинемия, галактозурия, протеинурия, гипераминоацидурия, накопление гликозилированного гемоглобина.
Патогенез. Избыток галактозы превращается в спирт галактитол (дульцитол), накапливающийся в хрусталике и осмотически привлекающий сюда воду. Изменяется солевой состав, нарушается конформация белков хрусталика, что приводит к катаракте в молодом возрасте. Катаракта возможна даже у плодов матерей с галактоземией, употреблявших молоко во время беременности.
При дефекте галактозо-1-фосфат-уридил-трансферазы АТФ постоянно расходуется на фосфорилирование галактозы и дефицит энергии угнетает активность многих ферментов, «токсически» действуя на нейроны, гепатоциты, нефроциты. Как результат возможны задержка психомоторного развития, умственная отсталость, некроз гепатоцитов и цирроз печени. В почках и кишечнике избыток галактозы и ее метаболитов ингибирует всасывание аминокислот.
Основы лечения. Исключение из рациона молока и других источников галактозы позволяет предотвратить развитие патологических симптомов. Однако сохранность интеллекта может быть достигнута только при ранней, не позднее первых 2 месяцев жизни, диагностике и вовремя начатом лечении.
Превращение фруктозы
В целом переход фруктозы в глюкозу осуществляется по двум направлениям. Сначала происходит активация фруктозы посредством фосфорилирования либо 6-го атома углерода при участии гексокиназы, либо 1-го атома при участии фруктокиназы.
В печени имеются оба фермента, однако гексокиназа имеет гораздо более низкое сродство к фруктозе и этот путь превращения слабо выражен. Образованный ею фруктозо-6-фосфат далее изомеризуется и глюкозо-6-фосфатаза отщепляет уже ненужный фосфат с получением глюкозы.
Если работает фруктокиназа, то образуется фруктозо-1-фосфат, под действием соответствующей альдолазы он превращается в глицеральдегид и диоксиацетонфосфат. Глицеральдегид фосфорилируется до глицеральдегидфосфата и вместе с диоксиацетонфосфатом они в дальнейших реакциях либо используются в гликолизе, либо в реакциях глюконеогенеза превращаются в фруктозо-6-фосфат и далее в глюкозу.
Особенностью мышц является отсутствие фруктокиназы, поэтому фруктоза в них превращается сразу в фруктозо-6-фосфат и поступает в реакции гликолиза или синтеза гликогена.
Пути метаболизма фруктозы и ее превращение в глюкозу
Особенностью метаболизма фруктозы является то, что фермент фруктокиназа является инсулин-независимым. В результате превращение фруктозы в пировиноградную кислоту и ацетил-SКоА происходит быстрее, чем для глюкозы. Это объясняется «игнорированием» лимитирующей реакции метаболизма глюкозы, катализируемой фосфофруктокиназой. Дальнейший метаболизм ацетил-SКоА в данном случае может привести к избыточному образованию жирных кислот и триацилглицеролов.Нарушения метаболизма фруктозы
Эссенциальная фруктозурия
Генетический дефект фруктокиназы приводит к доброкачественной эссенциальной фруктозурии, протекающей безо всяких отрицательных симптомов.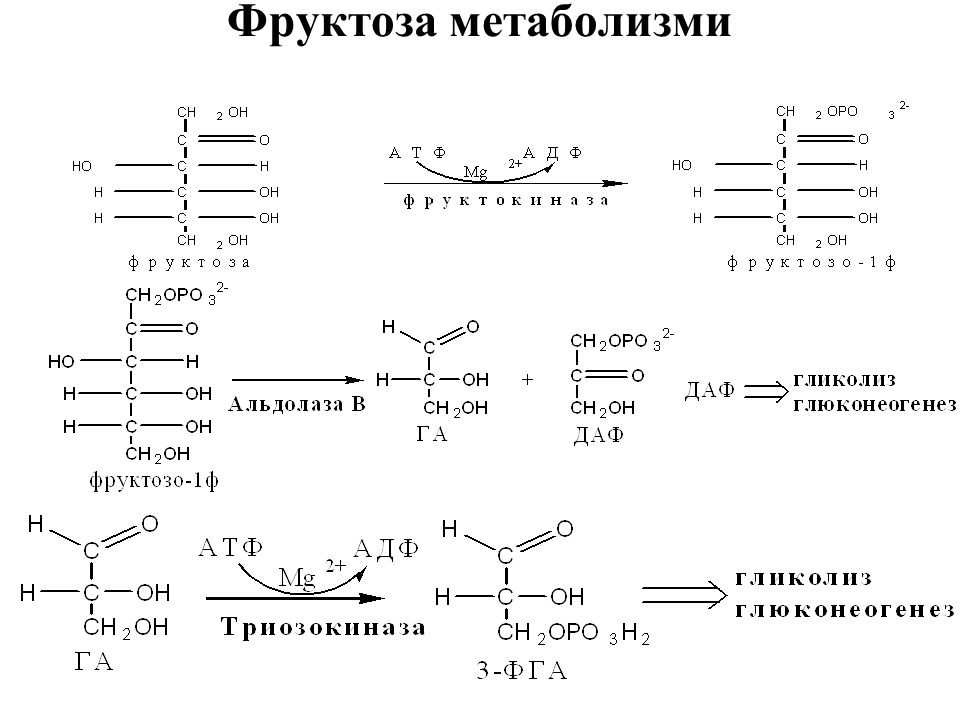
Наследственная фруктозурия
Заболевание формируется вследствие наследственных аутосомно-рецессивных дефектов других ферментов обмена фруктозы. Частота 1:20000.
Дефект фруктозо-1-фосфатальдолазы, которая в норме присутствует в печени, кишечнике и корковом веществе почек, проявляется после введения в рацион младенца соков и фруктов, содержащих фруктозу.
Патогенез связан со снижением мобилизации гликогена из-за ингибирования гликогенфосфорилазы фруктозо-1-фосфатом и ослаблением глюконеогенеза, т.к. дефектный фермент способен участвовать в реакциях аналогично фруктозо-1,6-дифосфат-альдолазе. Проявляется заболевание снижением концентрации фосфатов в крови, гиперфруктоземией, тяжелой гипогликемией. Отмечается вялость, нарушения сознания, почечный канальцевый ацидоз.
Диагноз ставится исходя из «непонятного» заболевания печени, гипофосфатемии, гиперурикемии, гипогликемии и фруктозурии. Для подтверждения проводят тест толерантности к фруктозе. Лечение включает диету с ограничением сладостей, фруктов, овощей.
Дефект фруктозо-1,6-дифосфатазы проявляется сходно с предыдущим, но не так тяжело.
Фруктоза и ожирение печени — почему сахар является ядом
Автор: доктор Джейсон Фанг
Фруктоза еще теснее связана с ожирением и диабетом, чем глюкоза. С точки зрения диетологии ни фруктоза, ни глюкоза не содержат жизненно необходимых питательных веществ. И в качестве подсластителей они похожи. Тем не менее, фруктоза приносит больший вред здоровью человека по сравнению с глюкозой из-за своего уникального метаболизма в организме.
Обмен веществ глюкозы и фруктозы отличается во многих отношениях. Почти каждая клетка в нашем организме может использовать глюкозу для получения энергии, но никакие клетки не умеют использовать фруктозу. Когда фруктоза попадает в наш организм, она может метаболизироваться только печенью. В то время как молекулы глюкозы могут рассеиваться по всему организму для использования в качестве энергии, молекулы фруктозы превращаются в управляемые ракеты, летящие в печень.
Когда мы едим много глюкозы, она начинает циркулировать практически в каждой клетке организма, которые помогают распределить эту нагрузку. Ткани тела, кроме печени, обрабатывают 80% съеденной глюкозы. Каждая клетка в организме, в том числе клетки сердца, легких, мышц, мозга и почек готовы полакомиться с глюкозного “шведского стола”. Это оставляет печени для переработки всего лишь 20% глюкозной нагрузки. Большая часть этой глюкозы превращается в гликоген для хранения, оставляя немного глюкозы в качестве субстрата для производства нового жира.
Чего нельзя сказать о фруктозе. Большие количества съеденного нутриента идут прямо в печень, потому что никакие другие клетки не могут помочь использовать или переработать ее, тем самым значительно увеличивая нагрузку на печень. Уровни углеводов и инсулина в ней могут быть в 10 раз выше, чем в других частях системы кровообращения организма. Таким образом, печень подвергается гораздо более высоким уровням углеводов – и фруктозы, и глюкозы – чем любой другой орган в нашем теле.
Сравните удар молотком и укол иглой: всё давление направлено на одну точку. Сахароза обеспечивает равные количества глюкозы и фруктозы. В то время, как целых 75 кг тканей обычного человека обрабатывают глюкозу, равное количество фруктозы должно быть отважно обработано только 2,2 кг печени. Что это означает на практике: фруктоза, возможно, имеет в 20 раз больше шансов вызвать ожирение печени (ключевая проблема ведущая к инсулинорезистентности) по сравнению с одной глюкозой. Это объясняет, каким образом многие примитивные сообщества могли переносить чрезвычайно высокоуглеводные диеты без развития гиперинсулинемии или резистентности к инсулину.
Печень преобразует фруктозу в глюкозу, лактозу и гликоген. Нет никаких ограничений в этой системе обмена веществ для фруктозы. Чем больше вы едите, тем больше вы обрабатываете. Когда ограниченные запасы гликогена заполнены, избыток фруктозы переходит непосредственно в жир печени через де-ново липогенез (DNL). Перекармливание фруктозой может увеличить DNL в 5 раз, а замена глюкозы на равный по калорийности объём фруктозы увеличивает объем жира в печени на 38% в течение всего 8 дней. Именно это ожирение печени имеет решающее значение для развития резистентности к инсулину.
Перекармливание фруктозой может увеличить DNL в 5 раз, а замена глюкозы на равный по калорийности объём фруктозы увеличивает объем жира в печени на 38% в течение всего 8 дней. Именно это ожирение печени имеет решающее значение для развития резистентности к инсулину.
Способности фруктозы вызывать ожирение печени уникальны среди углеводов. Ожирение печени непосредственно вызывает резистентность к инсулину, приводя в движение порочный круг: гиперинсулинемия – резистентность к инсулину. Кроме того, этот вредный эффект фруктозы не требует высоких уровней глюкозы или инсулина в крови, чтобы посеять хаос. Действуя через жировую болезнь печени и резистентность к инсулину, этот ведущий к ожирению эффект не заметен в краткосрочной перспективе – только на протяжении длительного времени.
Метаболизм этанола (спирта) весьма похож на метаболизм фруктозы. После проглатывания, ткани тела могут усвоить только 20% спирта, оставляя 80% для вывода непосредственно в печень, где он метаболизируется в ацетальдегид, стимулирующий де-ново липогенез. Суть заключается в том, что алкоголь просто превращается в печеночный жир.
Чрезмерное потребление этанола является хорошо известной причиной жировой дистрофии печени. Так как ожирение печени – это важный шаг на пути к резистентности к инсулину, неудивительно, что чрезмерное употребление этанола также является фактором риска развития метаболического синдрома.
Фруктоза и резистентность к инсулину
Тот факт, что переедание фруктозы может вызвать резистентность к инсулину, был известен еще в 1980 году. Здоровым участникам исследования давали избыточную фруктозу соответствующую 1000 калорий в день, и результаты показали ухудшение их чувствительности к инсулину на 25% – и это после 7 дней эксперимента! Тем, кому давали дополнительные 1000 калорий глюкозы в день, не показали подобного ухудшения.
Более позднее исследование 2009 года подтвердило, насколько легко фруктоза вызывает резистентность к инсулину у здоровых добровольцев.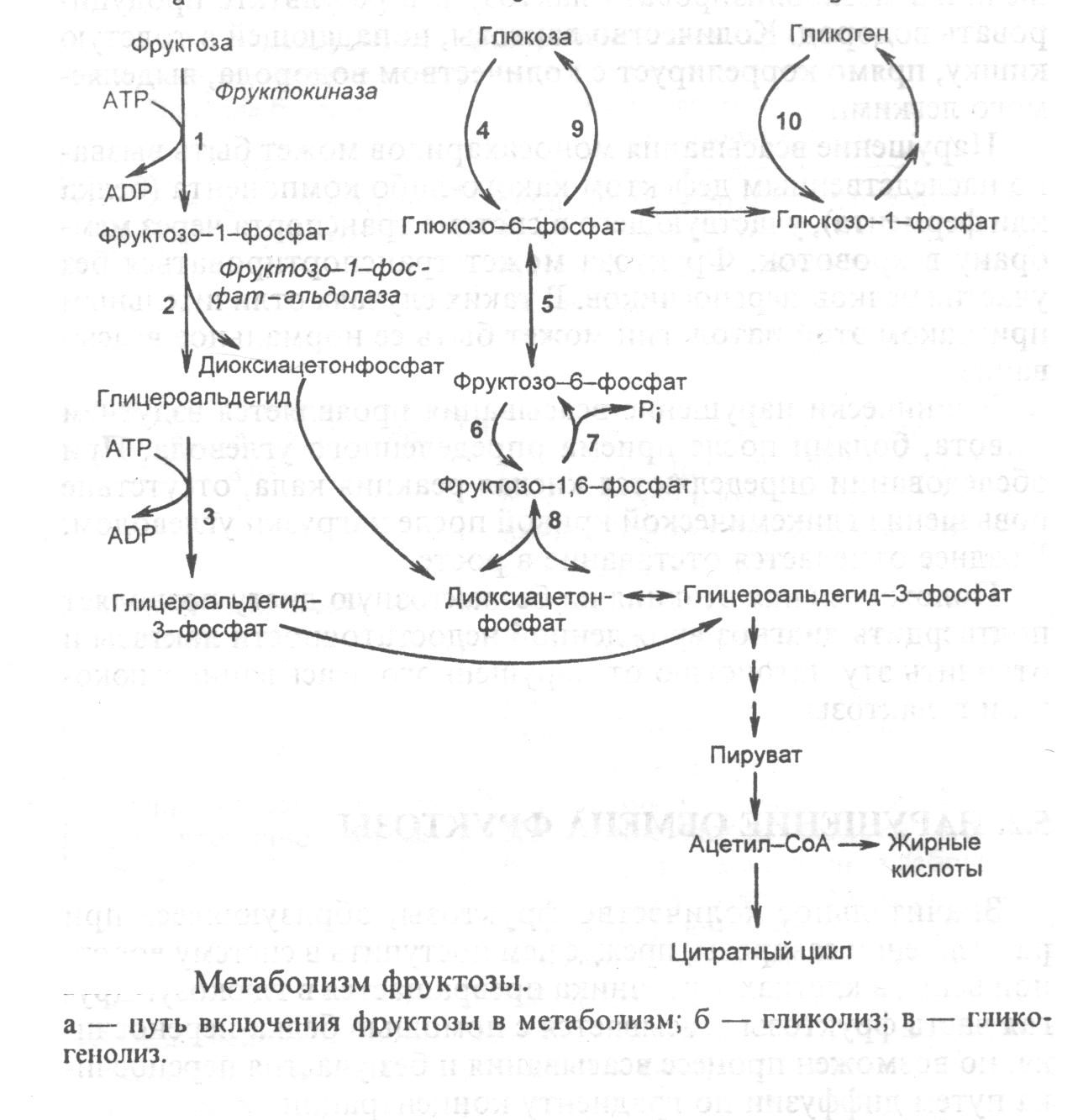 Испытуемым давали 25% от их ежедневных калорий в виде Kool-Aid (бренд ароматизированного напитка от компании Kraft Foods), подслащенного либо глюкозой, либо фруктозой. Хотя эти дозировки могут показаться высокими, многие сейчас потребляют примерно такой же объем сахара в своем рационе. В группе с фруктозой, в отличие от группы с глюкозой, увеличилась резистентность к инсулину так стремительно, что участники вполне могли быть продиагностированы как пред-диабетики. Ещё примечательнее другой факт: для развития подобного состояния понадобилось только 8 недель переедания фруктозы.
Испытуемым давали 25% от их ежедневных калорий в виде Kool-Aid (бренд ароматизированного напитка от компании Kraft Foods), подслащенного либо глюкозой, либо фруктозой. Хотя эти дозировки могут показаться высокими, многие сейчас потребляют примерно такой же объем сахара в своем рационе. В группе с фруктозой, в отличие от группы с глюкозой, увеличилась резистентность к инсулину так стремительно, что участники вполне могли быть продиагностированы как пред-диабетики. Ещё примечательнее другой факт: для развития подобного состояния понадобилось только 8 недель переедания фруктозы.
Всего 6 дней избытка фруктозы в вашем рационе вызывает резистентность к инсулину. 8 недель потребуется, чтобы получить пред-диабетическое состояние. Что же произойдет после десятилетий высокого потребления фруктозы? Результатом станет эпидемия сахарного диабета – то, что происходит сейчас с нами. Переедание фруктозы стимулирует ожирение печени и приводит непосредственно к резистентности к инсулину.
В переедании фруктозы действительно есть что-то зловещее. Да, доктор Роберт Ластиг прав. Сахар – это яд.
Факторы токсичности
Существует ряд причин, определяющих токсичность фруктозы. Во-первых, ее обмен веществ происходит исключительно в печени, поэтому практически весь ее объем после попадания в наш организм хранится в виде вновь созданного жира. В отличие от глюкозы, которую абсолютно все клетки могут метаболизировать.
Во-вторых, фруктоза преобразуется без ограничений. Чем больше мы потребляем фруктозы, тем интенсивнее происходит процесс печёночного де-ново липогенеза, и тем сильнее ожирение печени. Нет естественных тормозов в нашей системе, чтобы замедлить производство нового жира. Фруктоза непосредственно стимулирует DNL независимо от инсулина, поскольку диетическая фруктоза оказывает минимальное влияние на уровень глюкозы или инсулина в крови.
Метаболизм фруктозы регулируется менее жестко. Таким образом, он может перегрузить “экспортный механизм” печени, приведя к чрезмерному накоплению жира в ней.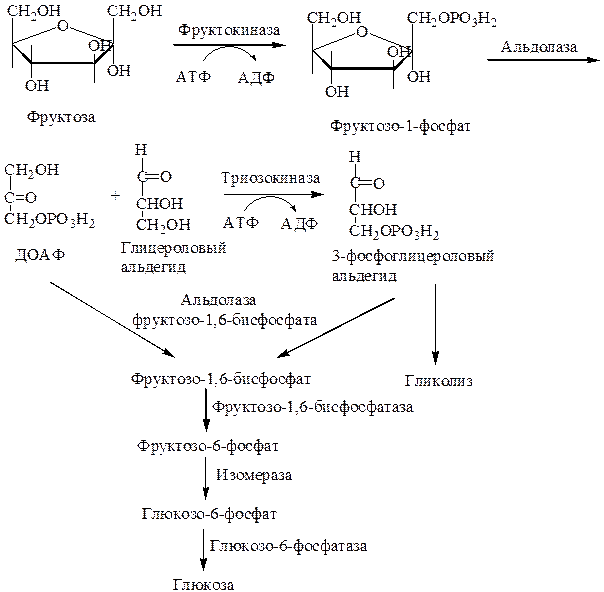 Мы поговорим о том, как печень пытается избавиться от вновь созданного жира в следующей главе.
Мы поговорим о том, как печень пытается избавиться от вновь созданного жира в следующей главе.
В-третьих, в организме не предусмотрено другой альтернативы для обработки фруктозы. Избыток глюкозы хранится безопасно и легко в печени в качестве гликогена. При необходимости гликоген трансформируется обратно в глюкозу для легкого доступа к энергии. Фруктоза не имеет еще одного механизма для удобного хранения. Она метаболизируется в жир, который не может легко быть преобразован обратно.
Поскольку фруктоза является натуральным сахаром, а также считается частью рациона человека с древнейших времен, мы должны всегда помнить первый принцип токсикологии. «Всё — яд, всё — лекарство; то и другое определяет доза». Тело имеет возможность обрабатывать небольшое количество фруктозы. Это не означает, что оно способно обрабатывать неограниченное ее количество без негативных последствий для здоровья.
Выводы
Фруктоза когда-то считались безвредной из-за своего низкого гликемического индекса. В краткосрочной перспективе, очевидных рисков для здоровья мало. Фруктоза проявляет свою токсичность в основном за счет долгосрочных эффектов, ведя к ожирению печени и резистентности к инсулину. Этот эффект часто измеряется десятилетиями, что приводит к немалому количеству дискуссий.
Таким образом, сахароза или высокофруктозный кукурузный сироп, примерно с равными частями глюкозы и фруктозы, играют двойную роль в процессе ожирения и развитии диабета 2 типа. Это не просто «пустые калории». Это нечто гораздо более зловещее, и люди постепенно начинают это понимать.
Глюкоза – рафинированный углевод, который напрямую стимулирует инсулин. Тем не менее, большая его часть может сжечься для получения энергии, оставив лишь небольшие количества продуктов обмена в печени. Тем не менее, очень высокие уровни потребления глюкозы также приводят к ожирению печени. Эффекты от потребления глюкозы очевидно проявляются в изменении ее уровня в крови и ответной реакции инсулина.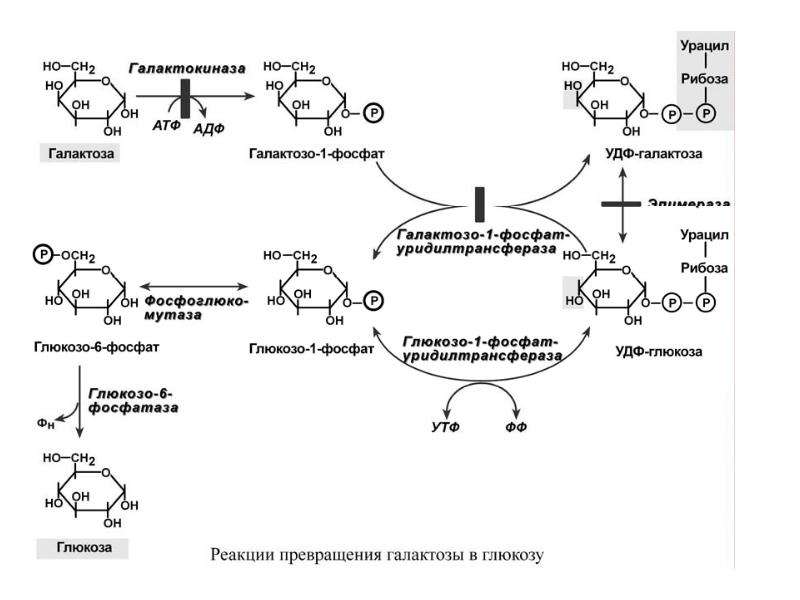
Переедание фруктозы производит жировое перерождение печени, что, в свою очередь, непосредственно создает резистентность к инсулину. Фруктоза на пять-десять раз чаще способна спровоцировать ожирение печени по сравнению с глюкозой, создавая зловещий порочный круг. Устойчивость к инсулину приводит к гиперинсулинемии, так как организм пытается «преодолеть» это сопротивление. Тем не менее, все это имеет неприятные последствия, потому что гиперинсулинемия усугубляется сопутствующей глюкозной нагрузкой, которая ведет к углублению состояния резистентности к инсулину.
Поэтому сахароза стимулирует выработку инсулина, как в краткосрочной, так и в долгосрочной перспективе. Таким образом, сахароза в разы более вредоносна, чем содержащие глюкозу крахмалы, такие, как амилопектин. Глядя на гликемический индекс, эффект от глюкозы очевиден, а эффект от фруктозы полностью скрыт. Этот факт уже давно ввел в заблуждение ученых, преуменьшавших роль сахара в эпидемии ожирения.
Воздействие инсулинорезистентности на ожирение накапливается в течение многих лет или даже десятилетий, прежде чем становится очевидным. Краткосрочные исследования полностью упускали этот эффект. Недавний системный анализ, рассмотревший множество исследований продолжительностью менее недели, пришёл к выводу, что фруктоза не вызывает особого эффекта, помимо содержащихся в ней калорий. Но последствия потребления фруктозы, например, ожирение, развиваются в течение десятилетий, а не недель. Если бы мы полагались на анализ только краткосрочных исследований о вреде курения, мы могли бы сделать ту же ошибку: посчитать, что курение не вызывает рак легких.
Сокращение в своем рационе сахара и сладостей всегда считается первым шагом к снижению веса практически во всех диетах на протяжении всей истории человечества. Сахароза – это не просто пустые калории или рафинированный углевод. Она гораздо опаснее, так как одновременно стимулирует и выработку инсулина и резистентность к нему. Наши предки всегда знали этот факт, даже если они не знали физиологии.
Мы попытались отрицать это на протяжение нашей 50-летней одержимости калориями. Пытаясь обвинить во всем избыточную калорийность, мы не распознали естественную опасность от переедания фруктозы. Но невозможно бесконечно отвергать истину, а за невежество приходится дорого платить. Мы поплатились двойной эпидемией: сахарного диабета 2 типа и ожирения. Но уникальные свойства сахара, ведущие к ожирению, были, наконец-то, снова признаны. Это была истина, котороая подавлялась на протяжение долгого времени.
Так что, когда доктор Ластиг представил свою лекцию в 2009 году и заявил, что сахар является ядом, весь мир слушал его с напряженным вниманием. Поскольку этот профессор эндокринологии говорил нам то, что мы уже инстинктивно знали и считали верным. Несмотря на все банальные возражения и заверения, что сахар не является проблемой, мир уже знал, в глубине души, настоящую правду. Сахар – это яд.
Оригинальная публикация на сайте dietdoctor.com
PS от LCHF.RU:
В российских магазинах продукты на фруктозе по-прежнему считаются диетическими и продаются в отделах здорового питания с соответствующей рекламой.
Здоровые низкоуглеводные продукты в нашем LCHF-магазине
Читайте также:
Сахар: горькая правда — главные тезисы легендарной лекции Роберта Ластига
Учёные объясняют почему сахар — это не еда
Почему нас тянет на сладкое, или сколько меда ели наши предки?
Новое исследование: мёд оказался не лучше сахара
Сироп агавы — натуральная, но нездоровая альтернатива сахару
Высокое потребление фруктозы во время беременности может повышать риск болезней среднего возраста у детей
Фруктоза усиливает тягу к вредной еде
Низкоуглеводная диета лечит ожирение печени, вызываемое… углеводами
Горькая правда о сладкой фруктозе / Здоровье / Независимая газета
В тучности обвиняют фруктозу.
Фото Интерпресс/PhotoXPress. ru
ru
Тучность – признанный спутник цивилизации. Количество карикатурно толстых молодых людей на улицах США ошеломляет. В Европе поменьше, но тоже много. Сейчас в мире тучных на 30% больше, чем недокормленных, чего никогда не бывало в истории. Заговорили уже об эпидемии тучности, охватившей западные страны, особенно среди детей. Речь не идет о красоте – речь идет о здоровье. Избыточный вес – одна из причин, вызывающих сердечно-сосудистые болезни и диабет.
Специалисты в области питания, особенно американские, долгое время считали причиной тучности потребление жиров, в первую очередь животных. Жиры начали решительно изгонять из всех продуктов. На полках супермаркетов появились обезжиренные сливки, обезжиренные сыры, обезжиренная сметана, даже обезжиренное сливочное масло. Однако людей с избыточным весом и всем букетом сопутствующих болезней меньше не стало.
Теперь американские эксперты пришли к выводу, что фактором, приводящим к ожирению, является сахар. В авторитетном научном журнале Nature они опубликовали статью под выразительным названием «Ядовитая правда о сахаре».
Один из авторов статьи – профессор Роберт Ластиг, педиатр и эндокринолог, глава Центра по борьбе с ожирением у детей и подростков при Калифорнийском университете в Сан-Франциско, уточняет, что имеется в виду не сахар как таковой, а сахар, который добавляется в безалкогольные напитки, полуфабрикаты, готовые кулинарные изделия.
За последние 50 лет потребление сахара в мире увеличилось втрое. Производители добавляют его во все мыслимые пищевые продукты. Эксперт в области диетологии и организации здравоохранения из Нью-Йоркского университета Мэрион Несле подчеркивает, что среднестатистический американец потребляет около четверти калорий именно с сахаром и зачастую не подозревает об этом.
Еще один автор статьи в журнале Nature – профессор Клэр Бриндис, педиатр, глава Центра глобальной репродуктивной медицины и директор Института по изучению политики в области здравоохранения при Калифорнийском университете в Сан-Франциско, говорит: «Если посмотреть на перечень ингредиентов продающегося в Америке хлеба, то и там все чаще обнаруживается сахар. В соусах, кетчупе, многих других продуктах раньше не было сахара, а сегодня он там есть. Избыточное присутствие сахаров характерно уже не только для лимонадов и прочих напитков такого рода, но и для многих других видов продовольствия».
В соусах, кетчупе, многих других продуктах раньше не было сахара, а сегодня он там есть. Избыточное присутствие сахаров характерно уже не только для лимонадов и прочих напитков такого рода, но и для многих других видов продовольствия».
Если раньше производители добавляли в продукты преимущественно сахарозу, то теперь ее все чаще заменяют фруктозой. Сахароза – это самый обычный сахар, тростниковый или свекловичный, он является дисахаридом, то есть состоит из двух моносахаридов – фруктозы и глюкозы. Попав в организм, сахароза быстро расщепляется на глюкозу и фруктозу. Фруктоза – самый сладкий из сахаров, в полтора раза слаще сахарозы и в три раза слаще глюкозы, добавлять ее выгоднее. Однако усваивается фруктоза совершенно иначе, чем глюкоза, которая является универсальным источником энергии для организма.
Фруктоза содержится практически во всех сладких ягодах и плодах, от нее, казалось бы, никакой опасности исходить не может. Но, как объясняет Роберт Ластиг, сахар, содержащийся во фруктах, потребляется вместе с растительными волокнами, которые, хоть сами в кишечнике и не усваиваются, регулируют процесс усвоения сахаров и тем самым уровень сахара в крови. Растительные волокна – своего рода противоядие, они препятствуют передозировке фруктозы в организме. А производители добавляют в свою продукцию фруктозу в чистом виде без сопутствующих балластных веществ.
Метаболизм фруктозы в организме очень отличается от метаболизма глюкозы и напоминает скорее метаболизм алкоголя, избыток фруктозы может вызывать недуги, типичные для алкоголизма: заболевания печени и сердечно-сосудистой системы. Фруктоза поступает прямиком в печень и может серьезно нарушить ее функцию, результатом чего часто становится метаболический синдром – чрезмерное увеличение массы висцерального жира, снижение чувствительности периферических тканей к инсулину, нарушение углеводного и липидного обмена, повышение артериального кровяного давления.
По оценкам профессора Ластига, сегодня уже три четверти всего бюджета здравоохранения США идет на лечение неинфекционных болезней – ожирения, сахарного диабета, рака, сердечно-сосудистых заболеваний, а в их развитии весьма заметную роль играет добавляемая в продукты питания фруктоза. По мнению американских ученых, фруктозу следует прежде всего исключить из перечня безопасных пищевых добавок. Это лишит промышленность права добавлять ее в любые продукты и в любом количестве.
По мнению американских ученых, фруктозу следует прежде всего исключить из перечня безопасных пищевых добавок. Это лишит промышленность права добавлять ее в любые продукты и в любом количестве.
Авторы статьи в Nature подчеркивают, что сахар в отношении воздействия на здоровье весьма схож с алкоголем: знакомство человека с ним неизбежно, он может вызывать зависимость (действует на «центр удовольствия» в мозге), токсичен (может непосредственно, даже без развития ожирения, стать причиной многих заболеваний), и избыточное потребление его несет негативные последствия для общества в целом. Поэтому оборот сахара необходимо законодательно регулировать так же, как оборот табака и алкоголя: ввести акцизный налог, ограничить продажу сладостей малолетним.
В России люди, как будто состоящие из жировых шаров, встречаются редко. Но тучных детей становится все больше. Роспотребнадзор запретил продавать в школьных буфетах пирожные и сладкую газировку. Однако бизнес у нас важнее детского здоровья. Запрет попросту игнорируют. Так что есть шансы догнать и перегнать Америку по количеству детей с ожирением.
Комментарии для элемента не найдены.
Потребление фруктозы и набор лишнего веса — Медицинский центр «Лотос»
Мальцева Оксана Александровна
Врач-диетолог
Учеными доказано: фруктоза способствует набору лишнего веса сразу несколькими способами:1. Фруктоза приводит к возникновению лептиновой резистентности. Лептин – это гормон жировой ткани, блокирующий чувство голода. Таким образом, чем больше человек потребляет фруктозы, тем меньше его организм становится невосприимчив к лептину. Именно поэтому он все время хочет есть, и, как правило, есть, быстро набирая лишние кг.
2. Чрезмерное потребление фруктозы также ведет к развитию инсулиновой резистентности, приводящей к ожирению печени, а затем к общему ожирению.
3. Фруктоза – причина сокращения выработки энергии организмом. Энергии становится меньше, как в печени, так и в мышцах. И именно с этим связан тот недостаток физической активности, который наблюдается у большинства людей в наши дни.
Люди мало и с неохотой двигаются не потому, что они ленивые, а потому, что им не хватает энергии. И каждое движение дается им с большим трудом.
Низкая физическая активность же, в свою очередь, ведет к тому, что масса тела увеличивается и двигаться становится все сложнее и сложнее.
Нехватка энергии ведет к набору лишнего веса не только за счет гиподинамии, а также потому, что фруктоза препятствует окислению жиров. А если жиры не окисляются, они накапливаются, а энергия не вырабатывается. Жировые отложения растут и не сгорают.
Фруктоза, как вид сахара, в больших объемах содержится в таких продуктах питания как: виноград, картофель, мед, земляника, яблоко, апельсины, бананы, арбуз, груша, черника, вишня.
Как видим, в любимых нами фруктах содержится максимальный объем фруктозы. Закономерный вопрос: сколько можно есть фруктозы и можно ли есть фрукты вообще?
Если вы уже страдаете резистентностью к инсулину и лептину, то не должны съедать в день больше 25 граммов фруктозы. А лучше – не более 15.
Понять, что у вас имеется резистентность к этим двум гормонам, довольно просто. Она есть у всех, кто имеет избыточную массу тела, у кого высокое артериальное давление, диабет или предиабетическое состояние.
Впрочем, по оценкам некоторых специалистов резистентность к инсулину и лептину имеет место у 4 из 5 взрослых людей. Так что, ограничить потребление фруктозы 25 граммами в сутки будет хорошо практически для всех.
Необходимо помнить: польза фруктов неоспорима, ведь в них большое количество питательных веществ, например, витаминов, а также антиоксидантов и клетчатки. Поэтому исключать их из своего меню не стоит. Однако во всем хороша мера. Запомните: фрукты – это десерт, точно такой же, как любая другая сладость. Так что, нельзя заканчивать обед десертом и фруктами. Либо фруктами, либо десертом. Впрочем, для тех, кто худеет, противопоказаны после обеда и фрукты, и десерт. А вот люди с нормальным весом должны выбрать что-то одно.
Так что, нельзя заканчивать обед десертом и фруктами. Либо фруктами, либо десертом. Впрочем, для тех, кто худеет, противопоказаны после обеда и фрукты, и десерт. А вот люди с нормальным весом должны выбрать что-то одно.
Если же вы уже страдаете лишним весом, вы должны быть крайне острожными с включением фруктов в свое меню. Употребляя фрукты, следует помнить, что вы едите фруктозу, и что ее безопасное ежедневное количество строго ограничено.
Каких фруктов стоит опасаться в большей мере, а какие безопасны для людей, борющихся с избыточным весом.
Первое, что нужно знать — вред или пользу фруктов не стоит измерять их калорийностью! Запомните: фруктоза низкокалорийна, но она намного опаснее больших калорий!
Конечно, не все плоды содержат избыточное количество фруктозы, существуют такие, которые можно назвать фруктами для похудения. Это – лимоны и лаймы, содержание фруктозы в которых минимально.
Грейпфруты, многие ягоды и киви также разрешено включать в меню при борьбе с лишним весом, так как содержание фруктозы в них не очень велико, а вот количество питательных веществ и антиоксидантов радует.
Однако, существуют фрукты, количество фруктозы в которых столь значительно, что их надо исключить из своего рациона тем, у кого есть малейшие признаки резистентности к инсулину и лептину – лишний вес, гипертония, и тем более диабет.
К таким опасным фруктам относят яблоки (особенно красные), абрикосы, персики, сливы.
Сухофрукты так же нельзя назвать чем-то безусловно полезным. В них много полезных веществ, например, микроэлементов, в частности калия. Но вот высокое содержание фруктозы заметно уменьшает их пользу.
Потому, если сравнивать сухофрукты со свежими фруктами, то предпочтение следует отдавать свежим. Сухие фрукты имеют более высокую концентрацию фруктозы и более низкое количество клетчатки.
Еще один вариант «безусловно полезного питания», с которым надо держать ухо востро, — это фруктовые соки. Пожалуй, это – самый плохой вариант употребления фруктов. Вся полезная клетчатка из них изымается. Выпить же сока можно и много, и очень много, нагрузив организм фруктозой выше всякой меры.
Вся полезная клетчатка из них изымается. Выпить же сока можно и много, и очень много, нагрузив организм фруктозой выше всякой меры.
Откровения тренера Виктора Васильева. Фруктоза: польза или вред?
Несмотря на то, что приближается самое страшное время для тренеров и диетологов — затяжной период тазиков оливье и прочих вкусных вредностей, сетевое издание «ВНовгороде.ру» продолжает вести здоровый образ жизни, чего желает и всем своим читателям. А сегодня в своей новой колонке Виктор Васильев разбирает важную и интересную для тех, кто в спорте, тему: фруктоза — польза или вред?
Слово «фруктоза» существует в сознании современного человека в тесной ассоциативной связи со словом фрукт, к которому сразу же прикрепляются расхожие штампы «польза», «витамины» «зож», дополненные рекламами соков, льющихся на нас с экранов масс-медиа, красивых стройных девушек кушающих яблоко на страницах глянцевых журналов. Разберемся, так ли это на самом деле!
Фруктоза принадлежит к классу простых углеводов, наряду с глюкозой. Однако, поскольку фруктоза является гораздо более сладкой, чем глюкоза, ее часть добавляют для подслащивания различных продуктов и блюд. Часто ее подменяют в составе кукурузным сиропом (который в свою очередь содержит от 65 до 85 % фруктозы).
Фруктоза получила такое широкое распространение в связи с её свойством не вызывать инсулиновый отклик, тем самым давая возможность людям, больным сахарным диабетом, побаловать себя сладким. Таким людям хочу напомнить, что решение о количестве принимаемой фруктозы в сутки должен принимать лечащий врач.
Как пачка фруктозы попадает в руки худеющей условно здоровой девушки или парня, которые наслушались доводов о её пользе, и пришли в магазин к витрине здорового питания? Ежедневное употребление чистой фруктозы в сочетании с употреблением продуктов и фруктов на завтрак, обед и ужин, даст организму человека существенное количество калорий. Но не дело не только в них…
Одной из эпидемий 21 века стал синдром Х или метаболический синдром, связанный с полной невосприимчивостью организма к инсулину. Ученые Калифорнийского университета, Центра по проблемам питания в Девисе, Пенсильванского университета выдвинули версию о вине фруктозы в развитии эпидемии ожирения, которое и вызывает этот метаболический синдром.
Ученые Калифорнийского университета, Центра по проблемам питания в Девисе, Пенсильванского университета выдвинули версию о вине фруктозы в развитии эпидемии ожирения, которое и вызывает этот метаболический синдром.
Оказывается, метаболизм фруктозы в нашем организме отличается от глюкозы. Попав в кровь, глюкоза, под воздействием инсулина, превращается в энергию, решая текущие энергетические потребности, или восполняет дефицит гликогена. Когда как фруктоза, расщепляясь в печени, или покрывает текущие энергетические потребности, или, если их нет, трансформируется в жир. Более того, избыток фруктозы в рационе приводит к тому, что печень выбрасывает больше триглицеридов в кровь, повышается количество липопротеинов низкой плотности, связанных с «плохим» холестерином. Согласно исследованиям, особенно это выражено у людей, уже страдающим избыточным весом. Повод задуматься о «безвредной» вкусняшке?
Но этого мало — фруктоза не просто трансформируется в жир. Она не стимулирует выработку гормонов, участвующих в энергетическом обмене – инсулина и лептина. Лептин вырабатывается в жировых клетках и является индикатором, который ограничивает запасы и производство жира, замедляя усвоение пищи и увеличивая расход энергии. Инсулин, в свою очередь, стимулирует выработку лептина. Вы приняли пищу (рациональный прием), сложные углеводы расщепились до глюкозы, организм выработал лептин и инсулин, чувство голода стало меньше. А теперь, кто сидел на фруктовых диетах и различных детокс программах, вспомните то чувство голода, что мучило вас всю диету? И это чувство голода будет гнать человека скушать еще больше «разрешенных» ему продуктов. Станет ли полезным перекус печеньем или зефиром на фруктозе? Или килограмма яблок (литра сока) вместо тарелки каши? Завтракать и ужинать стаканом сока? Задумайтесь, на самом ли деле так полезны активно пропагандируемые модные тенденции!
Значит ли это, что мы должны отказаться от употребления продуктов, содержащих фруктозу? Уверен, что нет.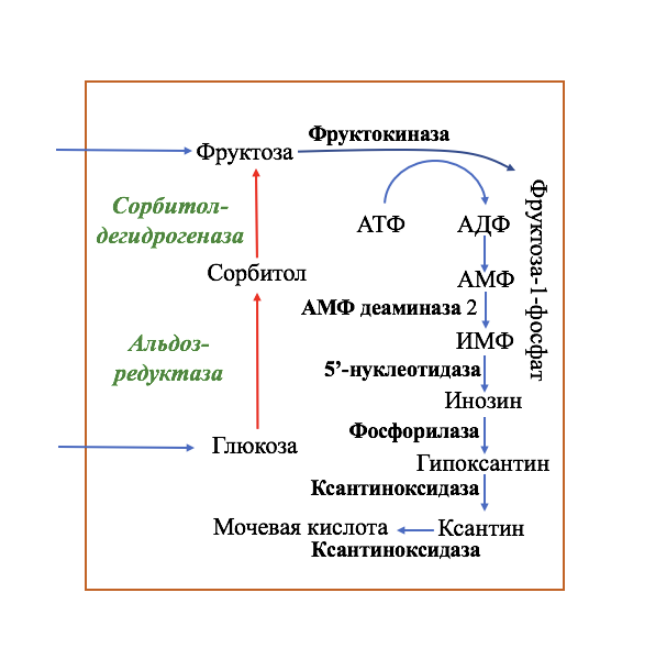 Исключая чистую фруктозу и кукурузный сироп, следует разумно употреблять в пищу продукты, богатые глюкозой: мед (39 грамм фруктозы на 100 грамм), сухофрукты (32 грамм на 100 грамм), варенье (30 грамм на 100 грамм), свежевыжатые соки (16 грамм на 200 мл), яблоки, вишня, виноград, киви, дыня (5-6 грамм на 100 грамм). Цифры небольшие, однако, представьте, если эти продукты станут основой питания и будут поглощаться килограммами? Средними показателями разумного потребления можно обозначить 15-20 грамм фруктозы в сутки. Для сравнения, в США, в рамках исследований, люди с диагностированным ожирением употребляли 90-120 гр фруктозы в сутки. Всякие же фруктозо-содержащие вкусности с полок «здорового» питания я бы советовал исключить полностью.
Исключая чистую фруктозу и кукурузный сироп, следует разумно употреблять в пищу продукты, богатые глюкозой: мед (39 грамм фруктозы на 100 грамм), сухофрукты (32 грамм на 100 грамм), варенье (30 грамм на 100 грамм), свежевыжатые соки (16 грамм на 200 мл), яблоки, вишня, виноград, киви, дыня (5-6 грамм на 100 грамм). Цифры небольшие, однако, представьте, если эти продукты станут основой питания и будут поглощаться килограммами? Средними показателями разумного потребления можно обозначить 15-20 грамм фруктозы в сутки. Для сравнения, в США, в рамках исследований, люди с диагностированным ожирением употребляли 90-120 гр фруктозы в сутки. Всякие же фруктозо-содержащие вкусности с полок «здорового» питания я бы советовал исключить полностью.
Желаю читателям придерживаться разумного подхода к питанию и, конечно же, добиться построения фигуры своей мечты!
Фото: pixabay
Метаболизм фруктозы и нарушение обмена веществ
J Clin Invest. 1 февраля 2018 г .; 128 (2): 545–555.
Сара А. Ханну
1 Отделение эндокринологии и метаболизма и Институт молекулярной физиологии Дьюка, Медицинский центр Университета Дьюка, Дарем, Северная Каролина, США.
Даниэль Э. Хаслам
2 Программа эпидемиологии питания, Жан Майер Центр исследований питания человека по проблемам старения Министерства сельского хозяйства США, Университет Тафтса, Бостон, Массачусетс, США.
Никола М. Маккеун
2 Программа эпидемиологии питания, Джин Майер, Центр исследований питания человека Министерства сельского хозяйства США, Университет Тафтса, Бостон, Массачусетс, США.
Марк А. Герман
1 Отделение эндокринологии и метаболизма и Институт молекулярной физиологии Дьюка, Медицинский центр Университета Дьюка, Дарем, Северная Каролина, США.
1 Отделение эндокринологии и метаболизма и Институт молекулярной физиологии Дьюка, Медицинский центр Университета Дьюка, Дарем, Северная Каролина, США.
2 Программа эпидемиологии питания, Жан Майер Центр исследований питания человека Министерства сельского хозяйства США по проблемам старения, Университет Тафтса, Бостон, Массачусетс, США.
Адрес для корреспонденции: Марк А. Херман, 300 Н. Дюк-стрит, Кармайкл Билдинг, Университет Дьюка, Дарем, Северная Каролина 27705, США. Телефон: 919.479.2378; Электронная почта: [email protected] Авторские права © 2018, Американское общество клинических исследований Эта статья цитируется в других статьях в PMC.Abstract
Повышенное потребление сахара все чаще считается одной из причин всемирных эпидемий ожирения и диабета и связанных с ними кардиометаболических рисков.В результате своих уникальных метаболических свойств фруктозный компонент сахара может быть особенно вредным. Диеты с высоким содержанием фруктозы могут быстро вызвать все ключевые признаки метаболического синдрома. Здесь мы рассматриваем биологию метаболизма фруктозы, а также потенциальные механизмы, с помощью которых чрезмерное потребление фруктозы может способствовать кардиометаболическому заболеванию.
Введение
Глюкоза является преобладающей формой циркулирующего сахара у животных, в то время как сахароза, дисахарид, состоящий из равных частей глюкозы и фруктозы, является преобладающим циркулирующим сахаром у растений.Поскольку растения составляют основу пищевой цепи, травоядные и всеядные животные хорошо приспособлены к использованию сахарозы для энергетических и биосинтетических нужд. Поскольку фруктоза не циркулирует в больших количествах у животных, съеденная фруктоза может быть уникальным образом расположена для передачи сигналов, связанных с потреблением сахара. Следовательно, понимание механизмов, с помощью которых ощущается фруктоза, может иметь важное значение для понимания адаптивной физиологии метаболизма сахарозы, а также потенциальных патофизиологических последствий чрезмерного потребления сахара.
Сахар в форме сахарозы или кукурузного сиропа с высоким содержанием фруктозы, оба из которых состоят из почти равных количеств глюкозы и фруктозы, добавляют во многие промышленные пищевые продукты. Подслащенные сахаром напитки (SSB) являются основным источником добавления сахара в рационы питания во всем мире и включают газированные напитки, напитки с фруктовым вкусом и спортивные напитки. В среднем SSB вносят примерно 7% дневных калорий (1) и почти 50% добавленных сахаров в рационе (2). Хотя тенденции в потреблении SSB в последние годы снизились, почти 66% молодежи США по-прежнему потребляют хотя бы один SSB в день (3).К другим основным факторам, влияющим на потребление сахара, относятся конфеты и десерты, на которые приходится от 4% до 9% дневной нормы потребления энергии в зависимости от возраста (2, 4).
Подслащенные сахаром напитки (SSB) являются основным источником добавления сахара в рационы питания во всем мире и включают газированные напитки, напитки с фруктовым вкусом и спортивные напитки. В среднем SSB вносят примерно 7% дневных калорий (1) и почти 50% добавленных сахаров в рационе (2). Хотя тенденции в потреблении SSB в последние годы снизились, почти 66% молодежи США по-прежнему потребляют хотя бы один SSB в день (3).К другим основным факторам, влияющим на потребление сахара, относятся конфеты и десерты, на которые приходится от 4% до 9% дневной нормы потребления энергии в зависимости от возраста (2, 4).
Вопрос о том, является ли повышенное потребление сахара основной причиной эпидемий ожирения, диабета 2 типа и неалкогольной жировой болезни печени, остается спорным (5–7). Хотя взаимосвязь между некоторыми показателями воздействия сахара в рационе и факторами кардиометаболического риска противоречива, большее потребление SSB неизменно ассоциируется с показателями более высокого кардиометаболического риска (5).Несколько крупных метаанализов связывают повышенное потребление SSB с увеличением массы тела, и большая часть, хотя и не все, этого увеличения веса, вероятно, связана с повышенным общим потреблением энергии (5, 8). SSB могут увеличивать кардиометаболический риск за счет увеличения висцерального ожирения, на которое приходится большая часть увеличения веса. Недавнее проспективное исследование показало, что у ежедневных потребителей SSB объем висцеральной жировой ткани за 6 лет увеличился на 29% по сравнению с непотребителями (9). Причинно-следственная связь подтверждается данными о том, что потребление 1 литра SSB ежедневно в течение 6 месяцев увеличивало висцеральный и печеночный жир, но не наблюдалось увеличения у тех, кто потреблял изокалорийное полубирочное молоко, некалорийную диетическую газировку или воду (10).Хотя повышенное висцеральное ожирение является основным кардиометаболическим фактором риска, SSB могут повышать риск независимо от ожирения. Например, ежедневное потребление SSB связано с нездоровым метаболическим профилем в разных слоях ИМТ и с повышенным риском диабета 2 типа независимо от ожирения (11, 12).
Например, ежедневное потребление SSB связано с нездоровым метаболическим профилем в разных слоях ИМТ и с повышенным риском диабета 2 типа независимо от ожирения (11, 12).
Гипертриглицеридемия — главный фактор риска сердечно-сосудистых заболеваний и еще один механизм, с помощью которого SSB могут увеличивать сердечно-сосудистый риск. Несколько крупных поперечных исследований изучали риск дислипидемии при приеме SSB, и эти исследования показывают, что распространенность дислипидемии увеличивается с увеличением потребления SSB (13, 14).Одно проспективное исследование показало, что употребление более 1 безалкогольного напитка в день увеличивает вероятность развития гипертриглицеридемии на 25% в течение 4 лет по сравнению с потреблением менее 1 безалкогольного напитка в день (15). Более того, два недавних проспективных когортных исследования показали, что ежедневное потребление SSB было связано примерно на 25% с повышенным риском развития ишемической болезни сердца как у мужчин, так и у женщин по сравнению с теми, кто не употреблял (13, 16).
Употребление SSB также связано с гипертонией, еще одним важным фактором риска сердечно-сосудистых заболеваний.Недавний метаанализ выявил умеренное увеличение риска гипертонии на 12% среди самых высоких потребителей SSB по сравнению с самыми низкими (17). Таким образом, прием SSB может способствовать развитию артериальной гипертензии, но может играть меньшую роль в этом факторе риска по сравнению с другими кардиометаболическими факторами риска.
На основании краткосрочных исследований перекармливания, проведенных преимущественно на животных, фруктозный компонент SSB и добавленный сахар оказались особенно вредными. Кормление животных большим количеством фруктозы может быстро вызвать множественные признаки метаболического синдрома, включая ожирение, дислипидемию, ожирение печени, гипертонию, инсулинорезистентность и диабет (18, 19).Некоторые, но не все краткосрочные исследования диетических вмешательств на людях также демонстрируют, что перекармливание фруктозы, но не глюкозы, может увеличить висцеральное ожирение, постпрандиальную гипертриглицеридемию и резистентность к инсулину, а влияние на определенные черты характера может зависеть от пола (20, 21). ). Одной из проблем таких исследований является то, что количество потребляемой фруктозы часто превышает количество, обычно обнаруживаемое в диетах ad libitum. Среднее потребление фруктозы среди населения США составляет примерно 9% от общего количества потребляемой энергии, в то время как потребители в 95-м процентиле в среднем составляют примерно 15% общей энергии из фруктозы (22).Напротив, многие интервенционные исследования непродолжительны (менее 4 недель) и включают потребление фруктозы с пищей, близкой к 25% от общего количества потребляемой энергии (23, 24).
). Одной из проблем таких исследований является то, что количество потребляемой фруктозы часто превышает количество, обычно обнаруживаемое в диетах ad libitum. Среднее потребление фруктозы среди населения США составляет примерно 9% от общего количества потребляемой энергии, в то время как потребители в 95-м процентиле в среднем составляют примерно 15% общей энергии из фруктозы (22).Напротив, многие интервенционные исследования непродолжительны (менее 4 недель) и включают потребление фруктозы с пищей, близкой к 25% от общего количества потребляемой энергии (23, 24).
Отсутствуют крупные рандомизированные контролируемые исследования диетических вмешательств, оценивающие влияние добавленных сахаров на кардиометаболические факторы риска в течение длительных периодов времени. Сложность, стоимость, соответствие и потенциальные этические проблемы, вероятно, препятствуют проведению таких исследований. Тем не менее, некоторые краткосрочные интервенционные исследования, даже те, которые находятся в диапазоне «нормального» потребления фруктозы, показывают, что фруктоза может быстро ухудшать промежуточные физиологические конечные точки, такие как циркулирующие липиды и чувствительность к инсулину у людей (25).В нескольких недавних обзорах всесторонне обсуждаются физиологические эффекты добавления фруктозы или сахара на патофизиологические конечные точки у людей (26, 27).
Понимание механизмов, с помощью которых изолированный моносахарид фруктозы может способствовать развитию метаболических заболеваний, может дать фундаментальное понимание патогенных механизмов, которые могут быть использованы для разработки новых диагностических, профилактических и терапевтических стратегий. Здесь мы рассмотрим биохимию и молекулярную генетику метаболизма фруктозы, а также потенциальные механизмы, с помощью которых чрезмерное потребление фруктозы способствует кардиометаболическим заболеваниям.Мы надеемся, что уроки, извлеченные из более глубокого понимания метаболизма фруктозы и кардиометаболического риска, вызванного фруктозой, могут быть применимы и к другим формам индуцированных диетой и генетически индуцированных метаболических заболеваний.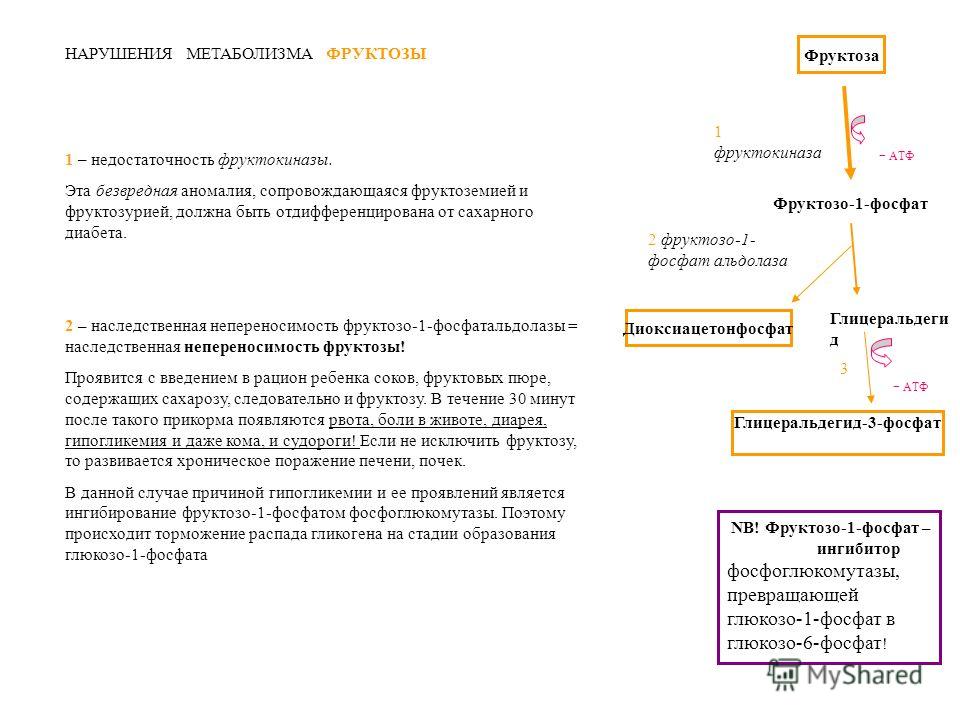
Абсорбция фруктозы
Проглоченная фруктоза в основном пассивно абсорбируется из просвета кишечника через транспортер гексозы SLC2A5, также известный как GLUT5, который имеет высокое сродство к фруктозе ( K m = 6 мМ). GLUT5 высоко экспрессируется на просветной мембране энтероцитов, а также базолатерально (28).Удаление Glut5 у мышей снижает всасывание фруктозы на 75% и вызывает расширение слепой и толстой кишки, а также накопление газа (29). Эти особенности указывают на мальабсорбцию фруктозы, которую часто называют причиной желудочно-кишечных симптомов у людей (30, 31). Способность кишечника абсорбировать фруктозу является насыщаемой (32), а способность здорового взрослого человека абсорбировать свободную фруктозу колеблется от менее 5 г до более 50 г (33). Неабсорбированная фруктоза может вызвать осмотическую нагрузку на дистальный отдел тонкой кишки и толстую кишку, что может способствовать развитию желудочно-кишечных симптомов (32).Более того, фруктоза может служить субстратом для бактериальной ферментации, что приводит к образованию газа и других бактериальных метаболитов, которые могут влиять на перистальтику кишечника и вызывать различные симптомы, такие как боль в животе и вздутие живота (34).
Кишечник GLUT5 Уровни мРНК и скорость транспорта фруктозы очень низкие пренатально и быстро увеличиваются с отлучением от груди независимо от диеты, но они могут быть дополнительно индуцированы после отлучения от диет, содержащих фруктозу (35). Недавние данные показали, что кормление с высоким содержанием фруктозы индуцирует белок, взаимодействующий с тиоредоксином кишечника (TXNIP), который связывает и регулирует опосредованный GLUT5 транспорт фруктозы в кишечнике (36).В соответствии с этим мы недавно показали, что белок, связывающий элемент, связывающий углеводы (ChREBP), фактор транскрипции, который реагирует на внутриклеточные углеводные питательные вещества и известный регулятор транскрипции TXNIP (37), также регулирует экспрессию GLUT5 в кишечнике и необходим для системной фруктозы. толерантность (38). В будущем будет интересно определить, влияет ли изменчивость экспрессии или функции GLUT5 или его регуляторных факторов на вариабельность всасывания фруктозы у людей.
толерантность (38). В будущем будет интересно определить, влияет ли изменчивость экспрессии или функции GLUT5 или его регуляторных факторов на вариабельность всасывания фруктозы у людей.
Промежуточный метаболизм фруктозы
Концентрация фруктозы в периферической плазме обычно составляет около 0,04 мМ, может резко увеличиваться в 10 раз после употребления фруктозы и возвращаться к уровню натощак в течение 2 часов (39–41). Этот быстрый клиренс в значительной степени опосредован эффективной экстракцией печенью. В то время как печень извлекает только 15–30% пероральной глюкозы, она способна извлекать 70% пероральной нагрузки фруктозой (42, 43). После приема фруктозы концентрация фруктозы в плазме может достигать низких миллимолярных концентраций в воротной вене, что сопровождается уровнем периферического кровообращения, равным примерно 0.2 мМ, что указывает на то, что периферические концентрации фруктозы редко превышают высокий микромолярный диапазон (44).
Транспортер глюкозы SLC2A2, также известный как GLUT2, имеет более низкое сродство к фруктозе ( K m = 11 мМ), чем GLUT5 (45). GLUT2 является второстепенным участником транспорта фруктозы в кишечнике (45), тогда как он, вероятно, является основным участником поглощения фруктозы печенью, поскольку GLUT5 не экспрессируется в печени в значительной степени (46, 47). SLC2A8, также известный как GLUT8, также может способствовать гепатоцеллюлярному транспорту фруктозы (48).Фруктоза — плохой субстрат для печеночной гексокиназы-глюкокиназы (GCK). Вместо этого кетогексокиназа (KHK, также известная как фруктокиназа) быстро фосфорилирует фруктозу с образованием фруктозо-1-фосфата (F1P). Высокая активность KHK и нечувствительность к энергетическому статусу клеток объясняют способность печени эффективно извлекать фруктозу. F1P метаболизируется до дигидроксиацетонфосфата (DHAP) и глицеральдегид-3-фосфата (G3P), которые входят в пулы гликолитических / глюконеогенных метаболитов ().
Биохимия фруктозы.
Попадая в гепатоциты, фруктоза фосфорилируется KHK до F1P. F1P расщепляется на DHAP и глицеральдегид с помощью ALDOB. Глицеральдегид фосфорилируется триозокиназой (TKFC, также известной как дигидроксиацетонкиназа 2 или DAK) с образованием гликолитического промежуточного глицеральдегид-3-фосфата (GA3P). И DHAP, и GA3P, производные фруктозы, входят в пул гликолитических / глюконеогенных метаболитов на уровне триозофосфата, и эти метаболиты имеют многочисленные метаболические судьбы. F1P также аллостерически регулирует метаболические ферменты (красные и зеленые линии), чтобы регулировать распределение производного фруктозы субстрата и других продуктов метаболизма, таких как мочевая кислота.AMPD3, аденозиндезаминаза; GA, глицеральдегид; IMP, монофосфат инозина; MTTP, микросомальный белок-переносчик триглицеридов; PYGL, гликогенфосфорилаза L; GYS2, гликогенсинтаза 2; PKLR, пируваткиназа, печень и эритроциты; PEP, фосфоенолпируват; ТАГ, триацилглицерин.
Клеточный метаболический статус и энергетический статус жестко регулируют стадию фосфофруктокиназы (PFK) в гликолизе, что ограничивает поток гликолита в печени (49). Напротив, производные фруктозы метаболиты попадают в триозофосфатный пул дистальнее PFK и, следовательно, обходят это ограничение.Поскольку печеночный фруктолиз неограничен, нагрузка фруктозой может привести к большим и быстрым расширениям гексозо- и триозофосфатных пулов, потенциально обеспечивая повышенный субстрат для всех центральных углеродных метаболических путей, включая гликолиз, гликогенез, глюконеогенез, липогенез и окислительное фосфорилирование.
Распределение углерода, полученного из фруктозы, среди основных метаболических путей зависит от общего нутритивного и эндокринного статуса животного, а также от статуса ключевых регулирующих контрольных точек в промежуточном метаболизме.Например, у голодных животных низкие уровни фруктозо-2,6-бифосфата ингибируют активность PFK и гликолиз, а также активируют фруктозо-1,6-бифосфатазу и производство глюкозы (50). Таким образом, у голодных животных триозофосфаты, полученные из фруктозы, предпочтительно проходят через глюконеогенный путь (51, 52). Судьба съеденной фруктозы также может зависеть от поступающих вместе питательных веществ. Например, введение физиологических концентраций фруктозы кормящим крысам и людям увеличивает уровни глюкозы и лактата в сыворотке, не влияя на накопление гликогена в печени (53, 54).Однако, когда фруктоза вводится с глюкозой, которая стимулирует секрецию инсулина, происходит заметное накопление гликогена (55). Хроническое потребление фруктозы может повлиять на программы экспрессии метаболических генов, что в дальнейшем влияет на распределение фруктозы. Эти механизмы будут описаны более подробно ниже.
Таким образом, у голодных животных триозофосфаты, полученные из фруктозы, предпочтительно проходят через глюконеогенный путь (51, 52). Судьба съеденной фруктозы также может зависеть от поступающих вместе питательных веществ. Например, введение физиологических концентраций фруктозы кормящим крысам и людям увеличивает уровни глюкозы и лактата в сыворотке, не влияя на накопление гликогена в печени (53, 54).Однако, когда фруктоза вводится с глюкозой, которая стимулирует секрецию инсулина, происходит заметное накопление гликогена (55). Хроническое потребление фруктозы может повлиять на программы экспрессии метаболических генов, что в дальнейшем влияет на распределение фруктозы. Эти механизмы будут описаны более подробно ниже.
Хотя печень метаболизирует большую часть съеденной фруктозы, сам кишечник может метаболизировать до 30% пероральной нагрузки фруктозы (56, 57). Все фруктолитические ферменты высоко экспрессируются в тонком кишечнике, особенно в тощей кишке, где наблюдаются самые высокие уровни GLUT5 (58).Подобно GLUT5, кишечная экспрессия фруктолитических и глюконеогенных ферментов, включая глюкозо-6-фосфатазу (G6PC), увеличивается при кормлении фруктозой (59) и зависит от активности GLUT5 и KHK (60). Однако большая часть прандиальной фруктозы не метаболизируется в кишечнике, а скорее проходит через воротную вену в печень (61, 62).
Помимо субстрата для метаболических процессов, метаболизм фруктозы в печени генерирует специфические метаболиты, которые также выполняют сигнальные функции ().Важно отметить, что F1P, специфический для фруктозы метаболит, продуцируемый KHK, оказывает сильный положительный регуляторный контроль GCK, способствуя его высвобождению из ингибирующего регуляторного белка GCK (GCKR). GCKR секвестрирует GCK в неактивном состоянии в ядре (63–65). «Каталитическое» количество фруктозы, частично за счет активации GCK, может способствовать усвоению и фосфорилированию глюкозы в печени, что приводит к быстрому накоплению гликогена (66). F1P может также усиливать синтез гликогена путем аллостерического ингибирования гликогенфосфорилазы (67, 68).Наконец, F1P также аллостерически активирует пируваткиназу, конечную стадию гликолиза, способствуя увеличению уровней циркулирующего лактата после приема фруктозы (69). В печени грызунов уровни F1P в печени увеличиваются в 10 раз до примерно 1 мМ в течение 10 минут после приема фруктозы и остаются повышенными в течение нескольких часов (70). Концентрации F1P только приблизительно 200 мкМ достаточны для ослабления ингибирующего действия GCKR на GCK (71). Таким образом, употребление фруктозы может иметь быстрое, надежное и устойчивое воздействие на усвоение глюкозы печенью и промежуточный метаболизм.
F1P может также усиливать синтез гликогена путем аллостерического ингибирования гликогенфосфорилазы (67, 68).Наконец, F1P также аллостерически активирует пируваткиназу, конечную стадию гликолиза, способствуя увеличению уровней циркулирующего лактата после приема фруктозы (69). В печени грызунов уровни F1P в печени увеличиваются в 10 раз до примерно 1 мМ в течение 10 минут после приема фруктозы и остаются повышенными в течение нескольких часов (70). Концентрации F1P только приблизительно 200 мкМ достаточны для ослабления ингибирующего действия GCKR на GCK (71). Таким образом, употребление фруктозы может иметь быстрое, надежное и устойчивое воздействие на усвоение глюкозы печенью и промежуточный метаболизм.
Метаболизм фруктозы активирует факторы транскрипции, включая ChREBP и SREBP1c и их коактиватор PGC1β, для координированной регуляции экспрессии генов метаболических ферментов, которые вносят вклад в фруктолиз, гликолиз, липогенез и производство глюкозы. Эти метаболические пути способствуют стеатозу, упаковке и секреции ЛПОНП, а также продукции глюкозы и образованию промежуточных липидов, которые могут влиять на чувствительность печени к инсулину и другие биологические процессы.АСАСА, ацетил-КоА карбоксилаза α; FASN, синтаза жирных кислот; ГПАТ, глицерин-3-фосфат-ацилтрансферазы; AGPAT, ацилглицерин-3-фосфатацилтрансфераза; DGAT, диацилглицерин ацилтрансфераза; ДАГ, диацилглицерин.
Хотя эффективность и скорость, с которой печень может извлекать и фосфорилировать проглоченную фруктозу, вероятно, важны для ее роли в интеграции пищевого и системного топливного метаболизма, этот устойчивый метаболизм также может иметь пагубные последствия. Например, уменьшение внутриклеточного свободного фосфата из-за быстрого фосфорилирования фруктозы в печени может увеличить выработку мочевой кислоты за счет активации АМФ-дезаминазы, что приводит к катаболизму АМФ в мочевую кислоту (72, 73).Кормление фруктозой может также стимулировать синтез пуринов, способствуя выработке мочевой кислоты (74). Повышенный уровень циркулирующей мочевой кислоты увеличивает риск подагры — состояния, характеризующегося болезненным воспалением из-за отложения кристаллов мочевой кислоты в суставах. Действительно, все больше данных указывает на то, что потребление сахара является фактором риска подагры (75). Более того, повышенный уровень мочевой кислоты в сыворотке и подагра связаны с другими кардиометаболическими факторами риска в различных популяциях (76–78). Значительный объем работ предполагает, что повышенный уровень мочевой кислоты может независимо регулировать важные аспекты метаболизма и способствовать кардиометаболическому риску (79–83).Однако исследования по менделевской рандомизации не подтверждают причинную роль циркулирующей мочевой кислоты в опосредовании кардиометаболических заболеваний (84). Связь между уровнями мочевой кислоты и кардиометаболическим риском может быть косвенной и может отражать активацию отдельных регулируемых фруктозой процессов, которые вносят вклад как в выработку мочевой кислоты, так и в кардиометаболический риск.
Повышенный уровень циркулирующей мочевой кислоты увеличивает риск подагры — состояния, характеризующегося болезненным воспалением из-за отложения кристаллов мочевой кислоты в суставах. Действительно, все больше данных указывает на то, что потребление сахара является фактором риска подагры (75). Более того, повышенный уровень мочевой кислоты в сыворотке и подагра связаны с другими кардиометаболическими факторами риска в различных популяциях (76–78). Значительный объем работ предполагает, что повышенный уровень мочевой кислоты может независимо регулировать важные аспекты метаболизма и способствовать кардиометаболическому риску (79–83).Однако исследования по менделевской рандомизации не подтверждают причинную роль циркулирующей мочевой кислоты в опосредовании кардиометаболических заболеваний (84). Связь между уровнями мочевой кислоты и кардиометаболическим риском может быть косвенной и может отражать активацию отдельных регулируемых фруктозой процессов, которые вносят вклад как в выработку мочевой кислоты, так и в кардиометаболический риск.
Печень находится на перекрестке метаболических путей и имеет решающее значение для измерения потребления питательных веществ и интеграции периферического статуса питательных веществ для регулирования системного хранения топлива по сравнению с обеспечением.В то время как гормоны, такие как инсулин и глюкагон, помогают информировать печень о системном топливном статусе, печень также хорошо настроена для интеграции сигналов, полученных непосредственно от топливных субстратов. В этом смысле сигнальные свойства F1P, полученного из фруктозы, и особенно его регуляция активности GCK, могут функционировать как развитый механизм, позволяющий печени использовать метаболизм фруктозы для «ощущения» сахара (например, сахарозы или кукурузного сиропа с высоким содержанием фруктозы). потребление. Устойчивая физиологическая активация печеночных GCK происходит только при потреблении фруктозосодержащих сахаров.Эта активация увеличивает чистое поглощение и хранение глюкозы в печени в виде гликогена и липидов. Интересно, что на супрафизиологическом / патологическом уровнях глюкоза сама может отделять GCK от GCKR и может вносить вклад в повышенную активность печеночных GCK, описанную у страдающих ожирением диабетиков и в генетических моделях ожирения и диабета (85, 86). Таким образом, в условиях неконтролируемого диабета печень может ошибочно воспринимать гипергликемию как состояние повышенного потребления сахара. Понимание метаболических эффектов печеночной «чувствительности к сахару» может иметь важное значение для понимания патофизиологии диабета и гипергликемии.
Генетические уроки метаболизма фруктозы
KHK существует в виде двух альтернативно сплайсированных изоформ, образующихся путем взаимного исключения соседних экзонов 3C и 3A в гене KHK (87, 88). Изоформа «А» экспрессируется повсеместно, но имеет низкую активность из-за относительно низкого сродства к ее субстрату ( K m = 8 мМ) (89). Экспрессия изоформы «C» в первую очередь ограничена метаболическими тканями, включая печень, почки и кишечник, и эта изоформа имеет гораздо более высокое сродство к фруктозе ( K m = 0.8 мМ) (89, 90). Мыши с дефицитом обеих изоформ были полностью защищены от метаболических заболеваний, вызванных фруктозой, даже несмотря на то, что уровни фруктозы в крови и моче были заметно повышены (91). Таким образом, повышенное содержание фруктозы в крови само по себе не вредно; скорее, метаболизм фруктозы важен для метаболического заболевания, вызванного фруктозой. Мутации с потерей функции в KHK вызывают у человека доброкачественное заболевание — эссенциальную фруктозурию, характеризующуюся нарушением метаболизма фруктозы в печени, что приводит к повышению уровня фруктозы в крови и моче после употребления сахарозы или фруктозы (92).Согласно наблюдениям на мышах, у людей с этим заболеванием не наблюдается никаких документально подтвержденных неблагоприятных последствий для здоровья. В целом, эти результаты предполагают, что ингибирование KHK может быть безопасной терапевтической стратегией для предотвращения метаболических заболеваний, вызванных фруктозой.
В отличие от глобальной делеции KHK, избирательная делеция изоформы A усугубляет неблагоприятные метаболические эффекты кормления фруктозой (91). Эти результаты предполагают две важные гипотезы: (а) метаболизм фруктозы за пределами тканей, которые экспрессируют изоформу C, не является незначительным и вносит вклад в клиренс фруктозы во всем организме, и (b) метаболизм фруктозы в тканях, экспрессирующих KHK-C, имеет решающее значение для фруктозы. -индуцированное нарушение обмена веществ.Это подтверждается недавними данными, показывающими, что селективный нокдаун KHK в печени мышей защищает от стеатоза, индуцированного фруктозой (93). Недавние данные также показывают, что измененный сплайсинг между изоформами KHK-A и KHK-C может способствовать развитию различных заболеваний, таких как гепатоцеллюлярная карцинома и сердечная недостаточность (94, 95).
Наследственная непереносимость фруктозы (HFI) — редкое аутосомно-рецессивное заболевание, вызванное дефицитом альдолазы B (ALDOB), которая сильно экспрессируется в печени, почках и тонком кишечнике (96).У людей с HFI возникают боли в животе, рвота, диарея, симптоматическая гипогликемия, гиперурикемия и, возможно, печеночная недостаточность и смерть после приема продуктов, содержащих фруктозу, сахарозу или сорбит (97). Точные механизмы, с помощью которых дефицит ALDOB вызывает симптомы, не совсем ясны. Модель мыши с дефицитом Aldob- имитирует состояние HFI человека (98). Эти мыши не могут развиваться и умирают, если их кормят пищей с высоким содержанием фруктозы. Интересно, что даже на диете без фруктозы у мышей с дефицитом Aldob- развивается стеатоз (98), возможно, из-за нарушения метаболизма эндогенно синтезированной фруктозы (99).
Производство эндогенной фруктозы
Хотя подавляющее большинство метаболизируемой фруктозы получают из пищевых источников сахара, животные, включая человека, способны синтезировать фруктозу эндогенно. Путь сорбитола (полиола), который активен в широком диапазоне тканей, отвечает за образование эндогенной фруктозы из глюкозы (100, 101). По этому пути глюкоза сначала восстанавливается до сорбита альдозоредуктазой (102). Затем сорбитол окисляется до фруктозы под действием сорбитолдегидрогеназы (103).Физиологически эндогенно синтезированная фруктоза является основным источником энергии для сперматозоидов и может иметь важное значение для фертильности (104–106). Плацента также может синтезировать сорбит, который развивающийся плод может использовать для синтеза фруктозы, что предполагает более широкую роль эндогенной фруктозы в репродуктивной биологии и биологии развития (107).
Активность пути сорбита увеличивается при диабетической гипергликемии (108). Синтез эндогенной фруктозы и метаболиты полиолов считаются ключевыми игроками в развитии микрососудистых осложнений диабета (109).Интересно, что концентрация фруктозы в сперме повышается при диабете 1 типа и при ожирении, при котором это связано с нарушением параметров спермы (105, 110). Только недавно было решено, может ли эндогенный синтез фруктозы происходить с достаточной скоростью, чтобы способствовать другим аспектам кардиометаболического риска, вызванного фруктозой. Глюкоза дозозависимо индуцирует альдолазоредуктазу в тканях человека, а хроническое воздействие диеты с высоким содержанием глюкозы индуцирует активацию пути полиола у мышей (99, 111).Это может быть механизмом, с помощью которого тяжелая гипергликемия может усугубить кардиометаболический риск. Кроме того, Lanaspa et al. сообщают, что производство эндогенной фруктозы и активация KHK в почках способствуют развитию диабетической нефропатии (112). Хотя сорбитолдегидрогеназа экспрессируется на высоком уровне в печени человека (113), требуется дальнейшее изучение того, является ли этот путь достаточно активным у человека, чтобы играть неблагоприятную метаболическую роль.
Влияние фруктозы на гомеостаз липидов
Как отмечалось выше, чрезмерное потребление фруктозы может оказывать значительное влияние на метаболизм липидов, способствуя как стеатозу, так и повышению уровня циркулирующих триглицеридов в форме липопротеинов очень низкой плотности (ЛПОНП).Накопление липидов в печени является результатом сочетания повышенного липогенеза в печени de novo (DNL), этерификации предварительно образованных жирных кислот, полученных из рациона или жировых запасов, снижения секреции ЛПОНП и снижения окисления жирных кислот в печени. Активация липогенной программы наблюдается сразу после однократной загрузки фруктозы и способствует увеличению секреции триглицеридов ЛПОНП (114, 115). Фруктоза также резко подавляет окисление жирных кислот печени (116). Таким образом, фруктоза способствует выработке триглицеридов в печени, обеспечивая как субстрат для синтеза жирных кислот и триглицеридов, так и активируя сигнальные системы для увеличения выработки липидов ().
Печень является основным участком DNL, процесса, посредством которого жирные кислоты синтезируются из пищевых предшественников, преимущественно углеводов (117). Из-за различий в метаболизме глюкозы и фруктозы в печени большая часть фруктозы, полученной с пищей, чем метаболитов глюкозы, доступна для преобразования в жир в печени через DNL у животных и людей (20, 118–120). Кроме того, метаболиты фруктозы, поступающие в триозофосфатный пул, находятся в равновесии с глицерин-3-фосфатом, который используется для синтеза глицеринового остова в триглицериде.Более того, метаболит малонил-КоА, генерируемый посредством DNL, ограничивает окисление жирных кислот, ингибируя карнитин-пальмитоилтрансферазу 1A (CPT1A), фермент, необходимый для транслокации жирных кислот в митохондрии (121). Ингибирование CPT1A дополнительно увеличивает доступность жирных кислот для производства триглицеридов. Триглицерид может включаться в липидные капли, приводя к стеатозу, или может быть включен в ЛОНП и секретироваться печенью.
Помимо обеспечения субстрата для липогенеза, хроническое потребление фруктозы увеличивает регуляцию транскрипции DNL за счет активации ключевых факторов транскрипции, включая белок 1c, связывающий регуляторный элемент стерола (SREBP1c) и белок, связывающий элемент, реагирующий на углеводы (ChREBP) (122).SREBP1c способствует синтезу липидов и регулируется на транскрипционном и посттрансляционном уровнях питательными веществами и гормонами. Инсулин является основным гормональным активатором печеночного SREBP1c (123, 124). Хотя острое кормление фруктозой напрямую не стимулирует секрецию инсулина, хроническое употребление фруктозы может привести к гиперинсулинемии, что может увеличить экспрессию и активацию SREBP1c в печени (125, 126). Фруктоза может также активировать SREBP1c независимо от инсулина, так как SREBP1c отвечает на кормление с высоким содержанием фруктозы у мышей с нокаутом печеночных рецепторов инсулина (LIRKO) (125).Потребление фруктозы также может способствовать стрессу ER, который может вызывать протеолитическое расщепление SREBP1c и липогенную программу (127, 128). Стресс ER, вызванный фруктозой, также может усиливать липогенез за счет активации фактора транскрипции x бокс-связывающего белка 1 независимо от других липогенных факторов транскрипции (129).
ChREBP связывает углеводные метаболиты с синтезом липидов, индуцируя ферменты, необходимые для DNL (130). ChREBP может также подавлять окисление жирных кислот за счет подавления ферментов, таких как CPT1A, частично за счет антагонизма рецептора α, активируемого пролифератором пероксисом (PPARα), ключевого регулятора транскрипции программы гена окисления жирных кислот (131, 132).ChREBP высоко экспрессируется в ключевых метаболических тканях, включая печень, жировую ткань, тонкий кишечник, островки поджелудочной железы и почки, где он регулирует углеводный обмен инсулино-независимым образом (37, 125, 130). Наблюдение, что мыши с дефицитом ChREBP не переносят диеты, содержащие фруктозу, но не диеты, содержащие декстрозу, предполагает особую роль ChREBP в регуляции метаболизма фруктозы (37, 133). Более того, активность ChREBP была заметно выше у крыс, получавших пищу с высоким содержанием фруктозы, по сравнению с изокалорийной диетой с высоким содержанием глюкозы (126).Недавно мы продемонстрировали, что прием фруктозы, но не глюкозы, резко и надежно индуцирует печеночную экспрессию мощной изоформы ChREBP-β вместе с ее липогенными, фруктолитическими и гликолитическими мишенями (133, 134). Механизм, с помощью которого метаболиты сахара активируют ChREBP, остается спорным, но включает аллостерическую активацию глюкозо-6-фосфатом, а также модуляцию другими углеводными метаболитами и посттрансляционные модификации (135–137). Нокдаун ChREBP с использованием антисмысловых олигонуклеотидов (ASO) у крыс, получавших фруктозу, снижал уровни циркулирующих триглицеридов и подтвердил роль ChREBP в опосредованной фруктозой дислипидемии, хотя стеатоз не изменился (138).В соответствии с этим, GWAS идентифицировал несколько общих SNP в локусе ChREBP , связанных с повышенным уровнем триглицеридов в сыворотке и низким уровнем холестерина ЛПВП (139, 140).
Избирательный эффект нокдауна ChREBP на циркулирующие триглицериды, но не на стеатоз в эксперименте, описанном выше, подчеркивает тот факт, что накопление жира в липидных каплях и секреция ЛПОНП являются разными процессами. ChREBP эффективно регулирует DNL, а DNL, индуцированная фруктозой, сильно коррелирует с индуцированной фруктозой гипертриглицеридемией (141).Однако у людей со стеатозом жирные кислоты, полученные из DNL, вносят незначительную долю жирных кислот в VLDL (142), и механистическая связь между DNL и секрецией VLDL остается неопределенной. Более того, ChREBP может оказывать влияние на увеличение циркулирующих триглицеридов независимо от увеличения секреции VLDL. ChREBP может трансактивировать экспрессию аполипопротеина APOC3, а также ангиопоэтин-подобного 8 (ANGPTL8), оба из которых могут ингибировать липопротеинлипазу и ограничивать клиренс VLDL (ссылки 143, 144 и).Таким образом, возможно, что кормление с высоким содержанием фруктозы может увеличить циркуляцию ЛПОНП как за счет увеличения продукции и секреции ЛПОНП, так и за счет уменьшения клиренса ЛПОНП, но точные механизмы еще предстоит определить.
Последствия чрезмерного потребления фруктозы.Метаболизм фруктозы в ключевых метаболических тканях, включая тонкий кишечник, печень и почки, может вносить вклад в различные кардиометаболические факторы риска, включая стеатоз, повышенное производство глюкозы, гипертриглицеридемию, повышенное ожирение и гипертензию.Фруктоза обеспечивает субстрат для метаболических процессов, которые способствуют кардиометаболическому риску, и задействует клеточные и гормональные сигнальные системы, которые регулируют эти метаболические и патологические процессы. LPL, липопротеинлипаза.
Коактиватор 1β PPARγ (PGC1β) представляет собой коактиватор транскрипции, который увеличивает активность нескольких ключевых факторов транскрипции, таких как PPARγ, PPARα, рецепторы, связанные с эстрогеном (ERR) и рецепторы печени X (LXR) (145, 146). PGC1β также может связывать SREBP1 и ChREBP и усиливать их транскрипционную активность (147, 148).ASO, нацеленные на PGC1β, предотвращают экспрессию SREBP1c и липогенез, что, в свою очередь, снижает накопление липидов в печени крыс, получавших фруктозу. ASO, нацеленные на PGC1β, также предотвращали увеличение ожирения, гликемии, инсулина и триглицеридов в плазме у крыс, получавших фруктозу. Таким образом, PGC1β имеет уникальную возможность координировать регуляцию активности как ChREBP, так и SREBP1c в контексте питания с высоким содержанием фруктозы.
Повышенное потребление сахара и фруктозы связано как с простым стеатозом, так и с прогрессированием более поздних форм неалкогольной жировой болезни печени (НАЖБП), включая неалкогольный стеатогепатит, фиброз и гепатоцеллюлярную карциному (149).Важные этапы синтеза DNL и VLDL происходят на мембране ER, а липогенез, индуцированный фруктозой, может вызывать стресс ER и стрессовую реакцию ER (150). Более того, сигнальные элементы в ответе на стресс ER могут вносить вклад в патогенез и прогрессирование НАЖБП (151). Недавняя работа Zhang et al. предполагает, что ChREBP может защищать печень от вызванного фруктозой стресса ER и воспаления печени (152). Однако недавно мы наблюдали, что у мышей с печеночно-специфическим ChREBP-нокаутом не развивается ER стресс или воспаление печени при введении диет с высоким содержанием фруктозы (38).Механизмы, с помощью которых фруктоза может способствовать прогрессированию НАЖБП, требуют дальнейшего изучения.
Влияние фруктозы на гомеостаз глюкозы
Фруктоза напрямую не стимулирует секрецию инсулина β-клетками поджелудочной железы (153, 154). Однако кормление с высоким содержанием фруктозы легко вызывает гиперинсулинемию на животных моделях. Более того, гиперинсулинемия более выражена у грызунов с высоким содержанием фруктозы по сравнению с кормлением с высоким содержанием декстрозы, несмотря на аналогичное увеличение массы тела и ожирения (155, 156).Точно так же кормление с гиперкалорией фруктозы увеличивает циркулирующий инсулин у людей (157). Гиперинсулинемия, вызванная фруктозой, которую часто считают признаком инсулинорезистентности, может быть результатом инсулинорезистентности в некоторой комбинации печени, мышц и / или жировой ткани.
Механизмы, с помощью которых кормление с высоким содержанием фруктозы вызывает гиперинсулинемию и инсулинорезистентность, остаются неясными. Вызванный фруктозой стеатоз может способствовать развитию инсулинорезистентности в печени за счет увеличения накопления диацилглицерина в печени, активации PKC и нарушения инсулино-опосредованной активации Akt2 (158–160).Однако вопрос о том, может ли стеатоз сам по себе вызывать резистентность к инсулину печени, остается спорным (131, 161). Помимо роли ChREBP в индуцированной фруктозой дислипидемии, Erion et al. продемонстрировали, что нокдаун ChREBP увеличивает периферическую чувствительность к инсулину у крыс, получавших много фруктозы (138). Неясно, было ли улучшение периферической чувствительности к инсулину напрямую связано с улучшением уровней циркулирующих липидов или ожирением. Недавно мы продемонстрировали, что, хотя печеночный ChREBP необходим для опосредованной фруктозой активации фруктолитических, гликолитических и липогенных ферментов, ChREBP также опосредует активацию G6PC, конечного фермента в производстве глюкозы (133).Мы показали, что индуцированное фруктозой, опосредованное ChREBP увеличение активности G6PC является основным детерминантом эндогенной продукции глюкозы. Более того, фруктоза активировала ChREBP и индуцировала G6PC в отсутствие FOXO1a, что указывает на то, что субстратная активация ChREBP и G6PC для увеличения продукции глюкозы преобладает над подавляющими эффектами инсулина (133). Эта ось передачи сигналов ChREBP / G6PC также консервативна у людей. Эти результаты согласуются с исследованиями диетических вмешательств на людях, показывающими, что либо эвкалорийное замещение, либо гиперкалорийное добавление фруктозы может иметь более значительное влияние на инсулинорезистентность печени, чем периферическая инсулинорезистентность (157).Однако, поскольку гиперинсулинемия сама по себе может вызывать периферическую инсулинорезистентность (162, 163), мы предполагаем, что хроническая гиперинсулинемия, которая компенсирует индуцированное фруктозой производство глюкозы, может впоследствии привести к периферической инсулинорезистентности. Эту гипотезу еще предстоит проверить экспериментально.
Влияние фруктозы на аппетит и ожирение
Повышенное потребление SSB и чрезмерное потребление фруктозы постоянно связаны с повышенным ожирением, что может быть связано с повышенным потреблением калорий, а также с влиянием на энергетический баланс и распределение питательных веществ, которые не зависят от количества потребляемых калорий.Фруктоза — один из самых сладких сахаров, а сладость обычно улучшает вкусовые качества пищи. Это, вероятно, способствует добавлению в пищу сахаров, содержащих фруктозу, таких как сахароза и кукурузный сироп с высоким содержанием фруктозы. Улучшение вкусовых качеств может улучшить пищевое поведение и, таким образом, способствовать перееданию (164). Более того, фруктоза и сахароза могут улучшить вкусовые качества и вызвать поведение, подобное зависимости, такое как переедание и зависимость, частично за счет стимуляции дофаминергических путей (165–168). В отличие от гедонической ценности фруктозы, также изучалось, влияет ли фруктоза на дополнительные сигнальные системы, регулирующие аппетит и пищевое поведение.Например, кормление с высоким содержанием фруктозы может вызвать резистентность к лептину, что, в свою очередь, может привести к увеличению потребления пищи и ожирению (169, 170). Кроме того, диетическая фруктоза снижает количество лептина по сравнению с изокалорийной диетической глюкозой, а фруктоза менее эффективна, чем глюкоза в подавлении орексигенного гормона грелина (171). У людей потребление фруктозы и глюкозы по-разному влияет на гипоталамический кровоток и реактивность коры головного мозга на пищевые сигналы, что предполагает возможность того, что фруктоза и глюкоза имеют разные эффекты на функцию мозга, которые могут влиять на пищевое поведение (172, 173).Механизмы, с помощью которых фруктоза и глюкоза по-разному регулируют аппетит и пищевое поведение, еще предстоит определить.
Перспективные исследования, в которых избыток фруктозы добавляется к привычному питанию, часто документируют спонтанное сокращение других форм потребления сахара, предлагая сильные механизмы обратной связи, которые конкретно регулируют потребление сахара (141). Такие компенсаторные механизмы могут затруднять точную оценку потребления сахара с пищей как в наблюдательных, так и в интервенционных исследованиях.FGF21 — это гормон печеночного происхождения, который регулирует гомеостаз энергии, глюкозы и липидов, а также может участвовать в механизме обратной связи, регулирующем выбор макроэлементов (ссылки 174–178 и). Повышенный уровень циркулирующего FGF21 связан с кардиометаболическими факторами риска, включая ожирение, НАЖБП, диабет 2 типа и инсулинорезистентность (179–181). FGF21 является мишенью транскрипции ChREBP (182), и прием фруктозы резко и надежно индуцирует циркулирующий FGF21, тогда как реакция на прием глюкозы менее существенна и задерживается (175).Циркулирующий FGF21, индуцированный фруктозой, может защищать печень от метаболических нарушений, вызванных фруктозой (183). Интересно, что данные на животных моделях предполагают, что индуцированный сахаром циркулирующий FGF21 может сигнализировать мозгу о подавлении дополнительного потребления сахара (184, 185). GWAS также поддерживает роль FGF21 в предпочтении макронутриентов, поскольку варианты в локусе FGF21 связаны с повышенным потреблением углеводов с пищей по сравнению с диетическими жирами в популяциях человека (186, 187). Однако варианты, связанные с повышенным потреблением углеводов, также связаны с повышенными уровнями циркулирующего FGF21, что несовместимо с моделью отрицательной обратной связи.Потребуются дополнительные исследования, чтобы понять роль FGF21 в контексте повышенного потребления сахара и фруктозы.
Влияние фруктозы на гипертонию
Механизмы, с помощью которых фруктоза способствует развитию гипертонии, менее изучены, чем ее влияние на гомеостаз глюкозы и липидов. Кормление грызунов с высоким содержанием фруктозы может увеличить всасывание соли в кишечнике отчасти за счет индукции кишечного анионообменника, Slc26a6 (188).Более того, эта индукция и связанная с ней гипертензия предотвращены у мышей с нокаутом по GLUT5 (188). Однако эти результаты противоречат тому факту, что мыши с нокаутом GLUT5 страдают генерализованной мальабсорбцией и заболевают при введении фруктозы. Джонсон и его коллеги предположили, что гиперурикемия, вызванная фруктозой, может нарушать функцию почек, что способствует развитию гипертонии (189). Однако, как обсуждалось выше, генетические данные не подтверждают основную роль гиперурикемии в кардиометаболическом заболевании.Поскольку фруктоза активно метаболизируется в почках, опосредованные фруктозой изменения в обработке солей в почках также могут быть важны. Однако это еще предстоит тщательно изучить и требует дальнейших исследований.
Выводы и направления на будущее
Сочетание механистических данных, подтверждающих роль чрезмерного употребления фруктозы, и эпидемиологических данных, подтверждающих роль SSB в развитии кардиометаболических заболеваний, поддерживает недавние диетические рекомендации по ограничению потребления сахара, опубликованные несколькими агентствами общественного здравоохранения, в том числе Американская кардиологическая ассоциация, Всемирная организация здравоохранения и Консультативный комитет по диетическим рекомендациям (2, 190, 191).Безопасные пороги потребления сахара и конкретные рекомендации по снижению кардиометаболического риска остаются предметом споров. Более того, реализация эффективных программ по изменению пищевых привычек остается сложной задачей. Однако первоначальные отчеты показывают, что «налоги на сахар» могут быть эффективными в сокращении потребления SSB (192, 193). Время покажет, могут ли такие подходы улучшить результаты в отношении здоровья. Надеюсь, что, улучшив наше понимание основных механизмов, с помощью которых сахар и фруктоза могут вызывать заболевания, мы сможем применить информированные и комплексные подходы к нашей текущей метаболической эпидемии.
Выражение признательности
Эта работа поддержана Американской кардиологической ассоциацией 16CSA285
(для MAH и NMM), NIH R01DK100425 (для MAH), NIH 5T32HL069772-15 (для DEH) и соглашением о Службе сельскохозяйственных исследований Министерства сельского хозяйства США 58-1950- 4-003 (в НММ).Изменения версии
Версия 1. 01.02.2018
Публикация для печати
Сноски
Конфликт интересов: M.A. Herman получил исследовательскую поддержку от Eli Lilly and Co.
Справочная информация: J Clin Invest. 2018; 128 (2): 545–555.
https://doi.org/10.1172/JCI96702.
Ссылки
1. Kit BK, Fakhouri TH, Park S, Nielsen SJ, Ogden CL. Тенденции потребления сахаросодержащих напитков среди молодежи и взрослых в США: 1999–2010 гг. Am J Clin Nutr. 2013. 98 (1): 180–188. DOI: 10.3945 / ajcn.112.057943. [PubMed] [CrossRef] [Google Scholar] 3. Rosinger A, Herrick K, Gahche J, Park S. Потребление сахаросодержащих напитков среди U.С. молодежь, 2011–2014 гг. Краткий обзор данных NCHS. 2017 (271): 1–8. [PubMed] [Google Scholar] 4. Вос М.Б. и др. Добавленный сахар и риск сердечно-сосудистых заболеваний у детей: научное заявление Американской кардиологической ассоциации. Тираж. 2017; 135 (19): e1017 – e1034. DOI: 10.1161 / CIR.0000000000000439. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 5. Хан Т.А., Сивенпайпер Ж.Л. Разногласия по поводу сахаров: результаты систематических обзоров и метаанализов ожирения, кардиометаболических заболеваний и диабета. Eur J Nutr.2016; 55 (приложение 2): 25–43. [Бесплатная статья PMC] [PubMed] [Google Scholar] 7. Stanhope KL. Потребление сахара, нарушение обмена веществ и ожирение: состояние противоречий. Критик Rev Clin Lab Sci. 2016; 53 (1): 52–67. DOI: 10.3109 / 10408363.2015.1084990. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 8. Малик В.С., Пан А., Уиллетт В.С., Ху Ф.Б. Сахарные напитки и увеличение веса у детей и взрослых: систематический обзор и метаанализ. Am J Clin Nutr. 2013. 98 (4): 1084–1102. DOI: 10.3945 / ajcn.113.058362. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 9. Ма Дж., МакКаун Н.М., Хван С.Дж., Хоффманн Ю., Жак П.Ф., Фокс С.С.. Потребление сахаросодержащих напитков связано с изменением висцеральной жировой ткани в течение 6 лет наблюдения. Тираж. 2016; 133 (4): 370–377. DOI: 10.1161 / CIRCULATIONAHA.115.018704. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 10. Маерск М. и др. Подслащенные сахарозой напитки увеличивают запасы жира в печени, мышцах и висцеральном жировом депо: 6-месячное рандомизированное интервенционное исследование.Am J Clin Nutr. 2012. 95 (2): 283–289. DOI: 10.3945 / ajcn.111.022533. [PubMed] [CrossRef] [Google Scholar] 11. Грин А. К., Жак П. Ф., Роджерс Г., Фокс С. С., Мейгс Дж. Б., МакКаун Н. М.. Сахарные напитки и распространенность метаболически аномального фенотипа в исследовании сердца Фрамингема. Ожирение (Серебряная весна) 2014; 22 (5): E157 – E163. DOI: 10.1002 / oby.20724. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 12. Имамура Ф. и др. Потребление сахаросодержащих напитков, искусственно подслащенных напитков и фруктовых соков и заболеваемость диабетом 2 типа: систематический обзор, метаанализ и оценка относительной доли населения.BMJ. 2015; 351: ч4576. [Бесплатная статья PMC] [PubMed] [Google Scholar] 13. де Конинг Л., Малик В.С., Келлог, доктор медицины, Римм Э.Б., Уиллетт В.С., Ху Ф.Б. Потребление сладких напитков, ишемическая болезнь сердца и биомаркеры риска у мужчин. Тираж. 2012. 125 (14): 1735–1741. DOI: 10.1161 / CIRCULATIONAHA.111.067017. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 14. Херт К.А., Фиск П.С., Ри Ю.С., Брант А.Р. Снижение потребления сахаросодержащих напитков улучшило отдельные биомаркеры риска хронических заболеваний среди взрослого населения США: с 1999 по 2010 год.Nutr Res. 2014; 34 (1): 58–65. DOI: 10.1016 / j.nutres.2013.10.005. [PubMed] [CrossRef] [Google Scholar] 15. Dhingra R, et al. Потребление безалкогольных напитков и риск развития кардиометаболических факторов риска и метаболического синдрома у взрослых среднего возраста в обществе. Тираж. 2007. 116 (5): 480–488. DOI: 10.1161 / CIRCULATIONAHA.107.689935. [PubMed] [CrossRef] [Google Scholar] 16. Фунг Т.Т., Малик В., Рексроде К.М., Мэнсон Дж. Э., Уиллетт В. К., Ху Ф. Б.. Потребление сладких напитков и риск ишемической болезни сердца у женщин.Am J Clin Nutr. 2009. 89 (4): 1037–1042. DOI: 10.3945 / ajcn.2008.27140. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 17. Джаялат В.Х. и др. Потребление сахаросодержащих напитков и эпизодическая гипертензия: систематический обзор и метаанализ предполагаемых когорт. Am J Clin Nutr. 2015; 102 (4): 914–921. DOI: 10.3945 / ajcn.115.107243. [PubMed] [CrossRef] [Google Scholar] 18. Ривен GM. Лекция Бантинга 1988. Роль инсулинорезистентности в заболеваниях человека. Сахарный диабет. 1988. 37 (12): 1595–1607. [PubMed] [Google Scholar] 19.Ривен GM. Инсулинорезистентность и компенсаторная гиперинсулинемия: роль в гипертонии, дислипидемии и ишемической болезни сердца. Am Heart J. 1991; 121 (4 pt 2): 1283–1288. [PubMed] [Google Scholar] 20. Стэнхоуп К.Л. и др. Употребление напитков, подслащенных фруктозой, а не глюкозой, увеличивает висцеральное ожирение и липиды, а также снижает чувствительность к инсулину у людей с избыточным весом / ожирением. J Clin Invest. 2009. 119 (5): 1322–1334. DOI: 10,1172 / JCI37385. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 21.Кузьма Ю.Н. и др. Нет различий в влиянии напитков, подслащенных фруктозой, кукурузным сиропом с высоким содержанием фруктозы или глюкозой, на системное воспаление или воспаление жировой ткани у взрослых с нормальным весом и ожирением: рандомизированное контролируемое исследование. Am J Clin Nutr. 2016; 104 (2): 306–314. DOI: 10.3945 / ajcn.115.129650. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 22. Marriott BP, Cole N, Lee E. Национальные оценки потребления фруктозы с пищей в США увеличились с 1977 по 2004 год. J Nutr. 2009; 139 (6): 1228С – 1235С.DOI: 10.3945 / jn.108.098277. [PubMed] [CrossRef] [Google Scholar] 23. Чунг М., Ма Дж., Патель К., Бергер С., Лау Дж., Лихтенштейн А. Х. Фруктоза, кукурузный сироп с высоким содержанием фруктозы, сахароза и неалкогольная жировая болезнь печени или показатели здоровья печени: систематический обзор и метаанализ. Am J Clin Nutr. 2014; 100 (3): 833–849. DOI: 10.3945 / ajcn.114.086314. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 24. Келишади Р., Мансуриан М., Хейдари-Бени М. Ассоциация потребления фруктозы и компонентов метаболического синдрома в исследованиях на людях: систематический обзор и метаанализ.Питание. 2014. 30 (5): 503–510. DOI: 10.1016 / j.nut.2013.08.014. [PubMed] [CrossRef] [Google Scholar] 25. Стэнхоуп К.Л. и др. Дозозависимое исследование потребления подслащенных напитков из кукурузного сиропа с высоким содержанием фруктозы на липидные / липопротеиновые факторы риска сердечно-сосудистых заболеваний у молодых людей. Am J Clin Nutr. 2015; 101 (6): 1144–1154. DOI: 10.3945 / ajcn.114.100461. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 26. Кампос В.К., Таппи Л. Физиологическая обработка диетических сахаров, содержащих фруктозу: последствия для здоровья.Int J Obes (Lond) 2016; 40 (приложение 1): S6 – S11. [PubMed] [Google Scholar] 27. Стэнхоуп К.Л., Шварц Дж.М., Гавел П.Дж. Неблагоприятные метаболические эффекты диетической фруктозы: результаты недавних эпидемиологических, клинических и механистических исследований. Curr Opin Lipidol. 2013. 24 (3): 198–206. DOI: 10.1097 / MOL.0b013e3283613bca. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 28. Патель К., Дуард В., Ю С, Гао Н., Феррарис Р.П. Регулирование всасывания фруктозы в кишечнике, зависящее от транспорта, метаболизма и эндосомного транспорта.FASEB J. 2015; 29 (9): 4046–4058. DOI: 10.1096 / fj.15-272195. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 29. Barone S, et al. Slc2a5 (Glut5) необходим для абсорбции фруктозы в кишечнике и возникновения гипертонии, вызванной фруктозой. J Biol Chem. 2009. 284 (8): 5056–5066. DOI: 10.1074 / jbc.M808128200. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 30. Гибсон ПР, Ньюнхэм Э., Барретт Дж. С., Шеперд С. Дж., Мьюир Дж. Дж. Обзорная статья: Мальабсорбция фруктозы и более широкая картина.Алимент Pharmacol Ther. 2007. 25 (4): 349–363. [PubMed] [Google Scholar] 31. Heizer WD, Southern S, McGovern S. Роль диеты в симптомах синдрома раздраженного кишечника у взрослых: обзорный обзор. J Am Diet Assoc. 2009. 109 (7): 1204–1214. DOI: 10.1016 / j.jada.2009.04.012. [PubMed] [CrossRef] [Google Scholar] 32. Rumessen JJ. Фруктоза и связанные с ней пищевые углеводы. Источники, потребление, абсорбция и клинические последствия. Сканд Дж Гастроэнтерол. 1992. 27 (10): 819–828. DOI: 10.3109 / 003655292048. [PubMed] [CrossRef] [Google Scholar] 33.Rumessen JJ, Gudmand-Høyer E. Способность абсорбции фруктозы у здоровых взрослых. Сравнение с сахарозой и входящими в ее состав моносахаридами. Кишечник. 1986. 27 (10): 1161–1168. DOI: 10.1136 / gut.27.10.1161. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 34. Скуг С.М., Бхаруча А.Е. Диетическая фруктоза и желудочно-кишечные симптомы: обзор. Am J Gastroenterol. 2004. 99 (10): 2046–2050. DOI: 10.1111 / j.1572-0241.2004.40266.x. [PubMed] [CrossRef] [Google Scholar] 37. Иидзука К., Брюик Р.К., Лян Дж., Хортон Дж. Д., Уеда К.Дефицит белка, связывающего элемент углеводного ответа (ChREBP), снижает липогенез, а также гликолиз. Proc Natl Acad Sci U S. A. 2004; 101 (19): 7281–7286. DOI: 10.1073 / pnas.0401516101. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 39. Сугимото К. и др. Снижение постпрандиальной гиперфруктоземии у людей с помощью экстракта листьев эвкалипта: рандомизированное двойное слепое плацебо-контролируемое перекрестное исследование. Food Sci Technol Res. 2010. 16 (5): 509–512. DOI: 10.3136 / fstr.16.509. [CrossRef] [Google Scholar] 40.Ваджуди П.Н., Паттерсон М.Э., Лим С., Йи Дж.К., Мао К.С., Ли В.Н. Измерение глюкозы и фруктозы в клинических образцах с помощью газовой хроматографии / масс-спектрометрии. Clin Biochem. 2010. 43 (1–2): 198–207. [Бесплатная статья PMC] [PubMed] [Google Scholar] 43. Tappy L, Lê KA. Способствует ли употребление фруктозы развитию неалкогольной жировой болезни печени? Клин Рес Гепатол Гастроэнтерол. 2012. 36 (6): 554–560. DOI: 10.1016 / j.clinre.2012.06.005. [PubMed] [CrossRef] [Google Scholar] 44. Сугимото К. и др. Экстракт листьев эвкалипта подавляет постпрандиальное повышение концентрации фруктозы в воротной, сердечной и периферической областях после приема сахарозы у крыс.J Clin Biochem Nutr. 2010. 46 (3): 205–211. DOI: 10.3164 / jcbn.09-93. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 45. Manolescu AR, Witkowska K, Kinnaird A, Cessford T, Cheeseman C. Облегченные переносчики гексозы: новые взгляды на форму и функции. Физиология (Bethesda) 2007; 22: 234–240. [PubMed] [Google Scholar] 46. Wood IS, Trayhurn P. Транспортеры глюкозы (GLUT и SGLT): расширенные семейства белков, транспортирующих сахар. Br J Nutr. 2003. 89 (1): 3–9. DOI: 10,1079 / BJN2002763. [PubMed] [CrossRef] [Google Scholar] 47.Карим С., Адамс Д.Х., Лалор П.Ф. Экспрессия в печени и клеточное распределение семейства переносчиков глюкозы. Мир Дж. Гастроэнтерол. 2012. 18 (46): 6771–6781. DOI: 10.3748 / wjg.v18.i46.6771. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 48. Дебош Б.Дж., Чен З., Сабен Д.Л., Финк Б.Н., Моли К.Х. Транспортер глюкозы 8 (GLUT8) опосредует индуцированный фруктозой липогенез de novo и макростеатоз. J Biol Chem. 2014. 289 (16): 10989–10998. DOI: 10.1074 / jbc.M113.527002. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 49.Boscá L, Corredor C. Является ли фосфофруктокиназа лимитирующей стадией гликолиза? Trends Biochem Sci. 1984. 9 (9): 372–373. DOI: 10.1016 / 0968-0004 (84)-7. [CrossRef] [Google Scholar] 51. Сестофт Л., Флерон П. Определение кинетических констант транспорта фруктозы и фосфорилирования в перфузированной печени крысы. Biochim Biophys Acta. 1974. 345 (1): 27–38. DOI: 10.1016 / 0005-2736 (74)
-9. [PubMed] [CrossRef] [Google Scholar] 52. Бьоркман О., Фелиг П. Роль почек в метаболизме фруктозы у людей, получающих голодание в течение 60 часов.Сахарный диабет. 1982; 31 (6 ч. 1): 516–520. [PubMed] [Google Scholar] 53. Сахебджами Х., Скалеттар Р. Эффекты инфузии фруктозы на метаболизм лактата и мочевой кислоты. Ланцет. 1971; 1 (7695): 366–369. [PubMed] [Google Scholar] 54. Берч Х. Б., Макс П., Гю К., Лоури Огайо. Промежуточные продукты метаболизма в печени крыс получают большое количество фруктозы или дигидроксиацетон. Biochem Biophys Res Commun. 1969. 34 (5): 619–626. DOI: 10.1016 / 0006-291X (69)
-9. [PubMed] [CrossRef] [Google Scholar] 55. Топпинг DL, Mayes PA. Сравнительные эффекты фруктозы и глюкозы на липидный и углеводный обмен перфузированной печени крысы.Br J Nutr. 1976; 36 (1): 113–126. DOI: 10,1079 / BJN19760062. [PubMed] [CrossRef] [Google Scholar] 56. Мавриас Д.А., Майер Р.Дж. Метаболизм фруктозы в тонком кишечнике. 1. Влияние кормления фруктозой на транспорт и метаболизм фруктозы в тонком кишечнике крыс. Biochim Biophys Acta. 1973; 291 (2): 531–537. DOI: 10.1016 / 0005-2736 (73)-X. [PubMed] [CrossRef] [Google Scholar] 57. Гинзбург V, Hers HG. О превращении фруктозы в глюкозу кишечником морской свинки. Biochim Biophys Acta. 1960; 38: 427–434.DOI: 10.1016 / 0006-3002 (60)-6. [PubMed] [CrossRef] [Google Scholar] 58. Рэнд Э.Б., Депаоли А.М., Дэвидсон Н.О., Белл Г.И., Бурант С.Ф. Последовательность, тканевое распределение и функциональная характеристика крысиного переносчика фруктозы GLUT5. Am J Physiol. 1993; 264 (6 пт 1): G1169 – G1176. [PubMed] [Google Scholar] 59. Cui XL, Сотеропулос П., Толиас П., Феррарис Р.П. Гены, чувствительные к фруктозе в тонком кишечнике новорожденных крыс. Physiol Genomics. 2004. 18 (2): 206–217. DOI: 10.1152 / Physiolgenomics.00056.2004. [PubMed] [CrossRef] [Google Scholar] 60.Патель К., Дуард В., Ю С., Тарабенджасин П., Гао Н., Феррарис Р.П. Индуцированное фруктозой увеличение экспрессии кишечных фруктолитических и глюконеогенных генов регулируется GLUT5 и KHK. Am J Physiol Regul Integr Comp Physiol. 2015; 309 (5): R499 – R509. DOI: 10.1152 / ajpregu.00128.2015. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 61. Bismut H, Hers HG, Van Schaftingen E. Превращение фруктозы в глюкозу в тонком кишечнике кролика. Переоценка прямого пути. Eur J Biochem. 1993. 213 (2): 721–726.DOI: 10.1111 / j.1432-1033.1993.tb17812.x. [PubMed] [CrossRef] [Google Scholar] 62. Холдсворт CD, изд. Сахар в питании . Нью-Йорк, Нью-Йорк, США: Raven Press; 1991. [Google Scholar] 63. Браун К.С., Калиновский С.С., Мегилл-младший, Дарем СК, Мухтиар К.А. Регуляторный белок глюкокиназы может взаимодействовать с глюкокиназой в ядре гепатоцита. Сахарный диабет. 1997. 46 (2): 179–186. [PubMed] [Google Scholar] 64. Никулеску Л., Вейга-да-Кунья М., Ван Шафтинген Э. Исследование механизма, с помощью которого фруктоза, гекситолы и другие соединения регулируют транслокацию глюкокиназы в гепатоцитах крысы.Биохим Дж. 1997; 321 (ч. 1): 239–246. [Бесплатная статья PMC] [PubMed] [Google Scholar] 65. Агиус Л. Глюкокиназа и молекулярные аспекты метаболизма гликогена в печени. Биохим Дж. 2008; 414 (1): 1–18. DOI: 10,1042 / BJ20080595. [PubMed] [CrossRef] [Google Scholar] 66. Макгиннесс О.П., Черрингтон А.Д. Влияние фруктозы на метаболизм глюкозы в печени. Curr Opin Clin Nutr Metab Care. 2003. 6 (4): 441–448. [PubMed] [Google Scholar] 67. Терстон Дж. Х., Джонс Э. М., Хаухарт РЭ. Снижение и угнетение гликогенфосфорилазы печени после фруктозы.Экспериментальная модель для изучения наследственной непереносимости фруктозы. Сахарный диабет. 1974. 23 (7): 597–604. DOI: 10.2337 / diab.23.7.597. [PubMed] [CrossRef] [Google Scholar] 68. Van Den Berghe G, Hue L, Hers HG. Влияние введения фруктозы на гликогенолитическое действие глюкагона. Исследование патогенеза наследственной непереносимости фруктозы. Биохим Дж. 1973; 134 (2): 637–645. DOI: 10,1042 / bj1340637. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 69. Эгглстон Л.В., Вудс ВЧ. Активация пируваткиназы печени фруктозо-1-фосфатом.FEBS Lett. 1970. 6 (1): 43–45. DOI: 10.1016 / 0014-5793 (70) 80038-2. [PubMed] [CrossRef] [Google Scholar] 70. Niewoehner CB, Gilboe DP, Nuttall GA, Nuttall FQ. Метаболические эффекты пероральной фруктозы в печени голодных крыс. Am J Physiol. 1984; 247 (4 п.1): E505 – E512. [PubMed] [Google Scholar] 71. Van Schaftingen E. Белок из печени крысы придает глюкокиназе свойство антагонистически регулироваться фруктозо-6-фосфатом и фруктозо-1-фосфатом. Eur J Biochem. 1989. 179 (1): 179–184. DOI: 10.1111 / j.1432-1033.1989.tb14538.x. [PubMed] [CrossRef] [Google Scholar] 72. Lanaspa MA, Tapia E, Soto V, Sautin Y, Sánchez-Lozada LG. Мочевая кислота и фруктоза: потенциальные биологические механизмы. Семин Нефрол. 2011. 31 (5): 426–432. DOI: 10.1016 / j.semnephrol.2011.08.006. [PubMed] [CrossRef] [Google Scholar] 73. ван ден Берге Г., Бронфман М., Ваннест Р., Херс Х. Г. Механизм истощения аденозинтрифосфата в печени после нагрузки фруктозой. Кинетическое исследование аденилатдезаминазы печени. Биохим Дж. 1977; 162 (3): 601–609.DOI: 10,1042 / bj1620601. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 74. Райвио К.О., Беккер М.А., Мейер Л.Дж., Грин М.Л., Нуки Г., Сигмиллер Дж.Э. Стимуляция синтеза пуринов человека de novo инфузией фруктозы. Metab Clin Exp. 1975. 24 (7): 861–869. DOI: 10.1016 / 0026-0495 (75)-X. [PubMed] [CrossRef] [Google Scholar] 75. Jamnik J, et al. Потребление фруктозы и риск подагры и гиперурикемии: систематический обзор и метаанализ проспективных когортных исследований. BMJ Open. 2016; 6 (10): e013191. DOI: 10.1136 / bmjopen-2016-013191. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 76. Chang CH, et al. Связь между гиперурикемией (HUC) и метаболическим синдромом (МС) у пожилых мужчин, находящихся в специализированных учреждениях. Arch Gerontol Geriatr. 2009; 49 (приложение 2): S46 – S49. [PubMed] [Google Scholar] 77. Лю М. и др. Связь между уровнем мочевой кислоты в сыворотке крови и метаболическим синдромом и его половыми различиями в пожилом населении китайской общины. Int J Endocrinol. 2014; 2014: 754678. [Бесплатная статья PMC] [PubMed] [Google Scholar] 78.Пуиг Дж. Г., Мартинес М. А.. Гиперурикемия, подагра и метаболический синдром. Curr Opin Rheumatol. 2008. 20 (2): 187–191. DOI: 10.1097 / BOR.0b013e3282f4b1ed. [PubMed] [CrossRef] [Google Scholar] 79. Lanaspa MA, et al. Мочевая кислота вызывает стеатоз печени, вызывая митохондриальный окислительный стресс: потенциальная роль в зависимой от фруктозы и независимой жировой ткани печени. J Biol Chem. 2012. 287 (48): 40732–40744. DOI: 10.1074 / jbc.M112.399899. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 80. Саутин Ю.Ю., Накагава Т., Жариков С., Джонсон Р.Дж.Побочные эффекты классического антиоксиданта мочевой кислоты в адипоцитах: окислительный / нитрозативный стресс, опосредованный НАДФН-оксидазой. Am J Physiol Cell Physiol. 2007; 293 (2): C584 – C596. DOI: 10.1152 / ajpcell.00600.2006. [PubMed] [CrossRef] [Google Scholar] 81. Tapia E, et al. Синергетический эффект блокады уриказы плюс физиологические количества фруктозы-глюкозы на гломерулярную гипертензию и окислительный стресс у крыс. Am J Physiol Renal Physiol. 2013; 304 (6): F727 – F736. DOI: 10.1152 / ajprenal.00485.2012. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 82.Choi YJ, et al. Мочевая кислота вызывает эндотелиальную дисфункцию из-за резистентности сосудов к инсулину, связанной с нарушением синтеза оксида азота. FASEB J. 2014; 28 (7): 3197–3204. DOI: 10.1096 / fj.13-247148. [PubMed] [CrossRef] [Google Scholar] 83. Lanaspa MA, et al. Мочевая кислота стимулирует фруктокиназу и ускоряет метаболизм фруктозы при развитии ожирения печени. PLoS One. 2012; 7 (10): e47948. DOI: 10.1371 / journal.pone.0047948. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 84. Ли X и др.Уровни мочевой кислоты в сыворотке и множественные исходы для здоровья: общий обзор данных обсервационных исследований, рандомизированных контролируемых испытаний и менделевских рандомизационных исследований. BMJ. 2017; 357: j2376. [Бесплатная статья PMC] [PubMed] [Google Scholar] 85. Бельфиоре Ф., Ромео Ф., Яннелло С., Саламон С. Соотношение глюкозо-6-фосфатаза / глюкокиназа в печени пациентов с ожирением и диабетом. Biochem Med Metab Biol. 1989. 41 (1): 77–80. DOI: 10.1016 / 0885-4505 (89)-X. [PubMed] [CrossRef] [Google Scholar] 86. Хуппонен Р., Карвонен И., Сотаниеми Э.Активность ферментов фосфорилирования глюкозы в печени и генерации НАДФН у крыс Zucker. Diabetes Res. 1989. 10 (3): 143–146. [PubMed] [Google Scholar] 87. Bonthron DT, Brady N, Donaldson IA, Steinmann B. Молекулярные основы эссенциальной фруктозурии: молекулярное клонирование и мутационный анализ кетогексокиназы (фруктокиназы) человека Hum Mol Genet. 1994. 3 (9): 1627–1631. DOI: 10,1093 / hmg / 3.9.1627. [PubMed] [CrossRef] [Google Scholar] 88. Hayward BE, Bonthron DT. Структура и альтернативный сплайсинг гена кетогексокиназы.Eur J Biochem. 1998. 257 (1): 85–91. DOI: 10.1046 / j.1432-1327.1998.2570085.x. [PubMed] [CrossRef] [Google Scholar] 89. Diggle CP и др. Кетогексокиназа: экспрессия и локализация основного фермента, метаболизирующего фруктозу. J Histochem Cytochem. 2009. 57 (8): 763–774. DOI: 10.1369 / jhc.2009.953190. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 90. Асипу А., Хейворд Б.Э., О’Рейли Дж., Бонтрон Д.Т. Свойства нормальных и мутантных рекомбинантных кетогексокиназ человека и их значение для патогенеза эссенциальной фруктозурии.Сахарный диабет. 2003. 52 (9): 2426–2432. DOI: 10.2337 / диабет.52.9.2426. [PubMed] [CrossRef] [Google Scholar] 91. Ишимото Т. и др. Противодействующие эффекты изоформ фруктокиназы C и A на метаболический синдром, индуцированный фруктозой, у мышей. Proc Natl Acad Sci U S. A. 2012; 109 (11): 4320–4325. DOI: 10.1073 / pnas.11199. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 92. Стейниц Х., Мизрахи О. Эссенциальная фруктозурия и наследственная непереносимость фруктозы. N Engl J Med. 1969; 280 (4): 222. [PubMed] [Google Scholar] 93.Softic S и др. Дивергентные эффекты глюкозы и фруктозы на липогенез в печени и передачу сигналов инсулина. J Clin Invest. 2017; 127 (11): 4059–4074. DOI: 10,1172 / JCI94585. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 95. Ли X и др. Переключение сплайсинга с кетогексокиназы-C на кетогексокиназу-A приводит к формированию гепатоцеллюлярной карциномы. Nat Cell Biol. 2016; 18 (5): 561–571. DOI: 10,1038 / ncb3338. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 96. Hers HG, Joassin G. [Аномалия печеночной альдолазы при непереносимости фруктозы] Enzymol Biol Clin (Базель) 1961; 1: 4–14.DOI: 10,1159 / 000458107. [PubMed] [CrossRef] [Google Scholar] 98. Oppelt SA, Sennott EM, Tolan DR. Нокаут альдолазы-B у мышей фенокопирует наследственную непереносимость фруктозы у людей. Mol Genet Metab. 2015; 114 (3): 445–450. DOI: 10.1016 / j.ymgme.2015.01.001. [PubMed] [CrossRef] [Google Scholar] 99. Lanaspa MA, et al. Производство и метаболизм эндогенной фруктозы в печени способствует развитию метаболического синдрома. Nat Commun. 2013; 4: 2434. [Бесплатная статья PMC] [PubMed] [Google Scholar] 100. Оутс П.Дж.Полиоловый путь и диабетическая периферическая нейропатия. Int Rev Neurobiol. 2002. 50: 325–392. [PubMed] [Google Scholar] 102. Ченг Х.М., Гонсалес Р.Г. Влияние высокого уровня глюкозы и окислительного стресса на метаболизм хрусталика, альдозоредуктазу и старческий катарактогенез. Metab Clin Exp. 1986; 35 (4 доп. 1): 10–14. [PubMed] [Google Scholar] 103. Jedziniak JA, Chylack LT, Jr, Cheng HM, Gillis MK, Kalustian AA и Tung WH. Путь сорбитола в хрусталике человека: альдозоредуктаза и полиолдегидрогеназа. Инвестируйте Ophthalmol Vis Sci.1981. 20 (3): 314–326. [PubMed] [Google Scholar] 104. Френетт Г., Табет М., Салливан Р. Путь полиола в придатке яичка и сперме человека. Дж. Андрол. 2006. 27 (2): 233–239. DOI: 10.2164 / jandrol.05108. [PubMed] [CrossRef] [Google Scholar] 105. Martini AC и др. Избыточный вес и семенные качества: исследование 794 пациентов. Fertil Steril. 2010. 94 (5): 1739–1743. DOI: 10.1016 / j.fertnstert.2009.11.017. [PubMed] [CrossRef] [Google Scholar] 106. Джаяраман В., Гош С., Сенгупта А., Шривастава С., Сонават Н. М., Нараян П. К.. Выявление биохимических различий между разными формами мужского бесплодия с помощью спектроскопии ядерного магнитного резонанса (ЯМР).J Assist Reprod Genet. 2014. 31 (9): 1195–1204. DOI: 10.1007 / s10815-014-0282-4. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 107. Hwang JJ и др. У беременных женщин уровень фруктозы в спинномозговой жидкости заметно повышен по сравнению с плазмой. PLoS One. 2015; 10 (6): e0128582. DOI: 10.1371 / journal.pone.0128582. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 108. Лоренци М. Путь полиола как механизм диабетической ретинопатии: привлекательный, неуловимый и устойчивый. Exp Diabetes Res.2007; 2007: 61038. [Бесплатная статья PMC] [PubMed] [Google Scholar] 109. Mathebula SD. Путь полиола: возможный механизм развития глазного диабета. Afr Vision Здоровье глаз. 2015; 74 (1): Ст. №13 [Google Scholar] 110. La Vignera S, Condorelli R, Vicari E, D’Agata R, Calogero AE. Сахарный диабет и параметры спермы. Дж. Андрол. 2012. 33 (2): 145–153. DOI: 10.2164 / jandrol.111.013193. [PubMed] [CrossRef] [Google Scholar] 111. Дас Б, Шривастава СК. Активация альдозоредуктазы из тканей человека. Сахарный диабет.1985. 34 (11): 1145–1151. [PubMed] [Google Scholar] 112. Lanaspa MA, et al. Производство эндогенной фруктозы и активация фруктокиназы опосредуют повреждение почек при диабетической нефропатии. J Am Soc Nephrol. 2014. 25 (11): 2526–2538. DOI: 10.1681 / ASN.2013080901. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 114. Sobrecases H, et al. Влияние кратковременного переедания фруктозой, жиром и фруктозой плюс жир на плазму и липиды печени у здоровых мужчин. Диабет Метаб. 2010. 36 (3): 244–246. DOI: 10.1016 / j.diabet.2010.03.003. [PubMed] [CrossRef] [Google Scholar] 115. Hudgins LC, Parker TS, Levine DM, Hellerstein MK. Двойной тест на липогенную чувствительность к пищевой фруктозе. J Clin Endocrinol Metab. 2011. 96 (3): 861–868. DOI: 10.1210 / jc.2010-2007. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 116. Топпинг DL, Mayes PA. Непосредственное воздействие инсулина и фруктозы на метаболизм перфузированной печени. Изменения секреции липопротеинов, окисления и этерификации жирных кислот, липогенеза и углеводного обмена.Biochem J. 1972, 126 (2): 295–311. DOI: 10,1042 / bj1260295. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 117. Hellerstein MK, Schwarz JM, Neese RA. Регуляция липогенеза de novo в печени у человека. Анну Рев Нутр. 1996; 16: 523–557. DOI: 10.1146 / annurev.nu.16.070196.002515. [PubMed] [CrossRef] [Google Scholar] 118. Паркс EJ, Skokan LE, Timlin MT, Dingfelder CS. Диетический сахар стимулирует синтез жирных кислот у взрослых. J Nutr. 2008. 138 (6): 1039–1046. [Бесплатная статья PMC] [PubMed] [Google Scholar] 119.Заварони I, Чен Ю.Д., Ривен ГМ. Изучение механизма индуцированной фруктозой гипертриглицеридемии у крыс. Metab Clin Exp. 1982; 31 (11): 1077–1083. DOI: 10.1016 / 0026-0495 (82)-X. [PubMed] [CrossRef] [Google Scholar] 120. Crescenzo R, Bianco F, Falcone I, Coppola P, Liverini G, Iossa S. Повышенный липогенез в печени de novo и митохондриальная эффективность в модели ожирения, вызванного диетами, богатыми фруктозой. Eur J Nutr. 2013. 52 (2): 537–545. DOI: 10.1007 / s00394-012-0356-у. [PubMed] [CrossRef] [Google Scholar] 121.МакГарри JD. Лекция Бантинга 2001: Нарушение регуляции метаболизма жирных кислот в этиологии диабета 2 типа. Сахарный диабет. 2002. 51 (1): 7–18. DOI: 10.2337 / диабет.51.1.7. [PubMed] [CrossRef] [Google Scholar] 122. Герман М.А., Самуэль В.Т. Сладкий путь к метаболической гибели: синтез фруктозы и липидов. Trends Endocrinol Metab. 2016; 27 (10): 719–730. DOI: 10.1016 / j.tem.2016.06.005. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 123. Ли С., Браун М.С., Гольдштейн Дж.Л. Бифуркация сигнального пути инсулина в печени крысы: mTORC1 необходим для стимуляции липогенеза, но не для ингибирования глюконеогенеза.Proc Natl Acad Sci U S. A. 2010; 107 (8): 3441–3446. DOI: 10.1073 / pnas.08107. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 125. Хаас Дж. Т. и др. Передача сигналов печеночного инсулина необходима для зависимой от ожирения экспрессии мРНК SREBP-1c, но не для зависимой от питания экспрессии. Cell Metab. 2012. 15 (6): 873–884. DOI: 10.1016 / j.cmet.2012.05.002. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 126. Ку HY, Miyashita M, Cho BH, Nakamura MT. Замена глюкозы в рационе фруктозой увеличивает активность ChREBP и белка SREBP-1 в ядре печени крысы.Biochem Biophys Res Commun. 2009. 390 (2): 285–289. DOI: 10.1016 / j.bbrc.2009.09.109. [PubMed] [CrossRef] [Google Scholar] 127. Каммун Х.Л. и др. Экспрессия GRP78 ингибирует инсулин и вызванную стрессом ER активацию SREBP-1c и снижает стеатоз печени у мышей. J Clin Invest. 2009. 119 (5): 1201–1215. DOI: 10.1172 / JCI37007. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 128. Flamment M, Hajduch E, Ferré P, Foufelle F. Новые взгляды на резистентность к инсулину, вызванную стрессом ER. Trends Endocrinol Metab.2012. 23 (8): 381–390. DOI: 10.1016 / j.tem.2012.06.003. [PubMed] [CrossRef] [Google Scholar] 129. Ли А.Х., Скапа Е.Ф., Коэн Д.Е., Глимчер Л.Х. Регуляция липогенеза печени с помощью фактора транскрипции XBP1. Наука. 2008. 320 (5882): 1492–1496. DOI: 10.1126 / science.1158042. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 130. Уеда К., Репа Дж. Дж. Белок, связывающий элемент углеводного ответа, ChREBP, фактор транскрипции, связывающий утилизацию глюкозы в печени и синтез липидов. Cell Metab. 2006. 4 (2): 107–110.DOI: 10.1016 / j.cmet.2006.06.008. [PubMed] [CrossRef] [Google Scholar] 131. Бенхамед Ф. и др. Липогенный фактор транскрипции ChREBP отделяет стеатоз печени от инсулинорезистентности у мышей и людей. J Clin Invest. 2012. 122 (6): 2176–2194. DOI: 10,1172 / JCI41636. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 132. Boergesen M, Poulsen Ll, Schmidt SF, Frigerio F, Maechler P, Mandrup S. ChREBP опосредует глюкозную репрессию экспрессии альфа-рецептора, активируемого пролифератором пероксисом, в бета-клетках поджелудочной железы.J Biol Chem. 2011. 286 (15): 13214–13225. DOI: 10.1074 / jbc.M110.215467. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 133. Ким М.С. и др. ChREBP регулирует индуцированное фруктозой производство глюкозы независимо от передачи сигналов инсулина. J Clin Invest. 2016; 126 (11): 4372–4386. DOI: 10,1172 / JCI81993. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 135. Dentin R, et al. Глюкозо-6-фосфат, а не ксилулозо-5-фосфат, необходим для активации ChREBP в ответ на глюкозу в печени.J Hepatol. 2012. 56 (1): 199–209. DOI: 10.1016 / j.jhep.2011.07.019. [PubMed] [CrossRef] [Google Scholar] 136. Filhoulaud G, Guilmeau S, Dentin R, Girard J, Postic C. Новое понимание регуляции и функции ChREBP. Trends Endocrinol Metab. 2013. 24 (5): 257–268. DOI: 10.1016 / j.tem.2013.01.003. [PubMed] [CrossRef] [Google Scholar] 137. Baraille F, Planchais J, Dentin R, Guilmeau S, Postic C. Интеграция ChREBP-опосредованного восприятия глюкозы в метаболизм всего тела. Физиология (Bethesda) 2015; 30 (6): 428–437.[PubMed] [Google Scholar] 138. Эрион Д.М. и др. Роль связывающего элемент углеводного ответа белка у самцов крыс, получавших фруктозу. Эндокринология. 2013. 154 (1): 36–44. DOI: 10.1210 / en.2012-1725. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 139. Кунер Дж. С. и др. Полногеномное сканирование выявляет вариации MLXIPL, связанные с триглицеридами плазмы. Нат Жене. 2008. 40 (2): 149–151. DOI: 10.1038 / нг.2007.61. [PubMed] [CrossRef] [Google Scholar] 140. Kathiresan S, et al. Шесть новых локусов, связанных с холестерином липопротеинов низкой плотности крови, холестерином липопротеинов высокой плотности или триглицеридами у людей.Нат Жене. 2008. 40 (2): 189–197. DOI: 10,1038 / нг.75. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 141. Taskinen MR, et al. Неблагоприятные эффекты фруктозы на кардиометаболические факторы риска и метаболизм липидов в печени у пациентов с абдоминальным ожирением. J Intern Med. 2017; 282 (2): 187–201. DOI: 10.1111 / joim.12632. [PubMed] [CrossRef] [Google Scholar] 142. Доннелли К.Л., Смит К.И., Шварценберг С.Дж., Джессурун Дж., Болдт, доктор медицины, Паркс Е.Дж. Источники жирных кислот, хранящиеся в печени и секретируемые липопротеинами у пациентов с неалкогольной жировой болезнью печени.J Clin Invest. 2005. 115 (5): 1343–1351. DOI: 10,1172 / JCI23621. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 143. Карон С. и др. Активация транскрипции экспрессии аполипопротеина CIII глюкозой может способствовать диабетической дислипидемии. Артериосклер Thromb Vasc Biol. 2011; 31 (3): 513–519. DOI: 10.1161 / ATVBAHA.110.220723. [PubMed] [CrossRef] [Google Scholar] 144. Fu Z, Berhane F, Fite A, Seyoum B, Abou-Samra AB, Zhang R. Повышенный уровень циркулирующего липазина / бетатрофина при диабете 2 типа и ожирении у человека.Научный отчет 2014; 4: 5013. [Бесплатная статья PMC] [PubMed] [Google Scholar] 145. Лин Дж., Пуигсервер П., Донован Дж., Тарр П., Шпигельман Б.М. Коактиватор гамма рецептора, активируемого пролифератором пероксисом, 1beta (PGC-1beta), новый коактиватор транскрипции, связанный с PGC-1, связанный с фактором клетки-хозяина. J Biol Chem. 2002. 277 (3): 1645–1648. DOI: 10.1074 / jbc.C100631200. [PubMed] [CrossRef] [Google Scholar] 146. Lin J, Handschin C, Spiegelman BM. Метаболический контроль с помощью семейства коактиваторов транскрипции PGC-1.Cell Metab. 2005. 1 (6): 361–370. DOI: 10.1016 / j.cmet.2005.05.004. [PubMed] [CrossRef] [Google Scholar] 147. Lin J, et al. Гиперлипидемические эффекты пищевых насыщенных жиров, опосредованные посредством коактивации SREBP PGC-1β. Клетка. 2005. 120 (2): 261–273. DOI: 10.1016 / j.cell.2004.11.043. [PubMed] [CrossRef] [Google Scholar] 148. Chambers KT, et al. PGC-1β и ChREBP участвуют в совместной регуляции липогенеза печени в зависимости от концентрации глюкозы. Mol Metab. 2013. 2 (3): 194–204. DOI: 10.1016 / j.молмет.2013.05.001. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 149. Лим Дж. С., Митус-Снайдер М., Валенте А., Шварц Дж. М., Лустиг Р. Х. Роль фруктозы в патогенезе НАЖБП и метаболического синдрома. Нат Рев Гастроэнтерол Гепатол. 2010. 7 (5): 251–264. DOI: 10.1038 / nrgastro.2010.41. [PubMed] [CrossRef] [Google Scholar] 150. Чан С.М. и др. Активация PPARα улучшает инсулинорезистентность печени и стеатоз у мышей, получавших много фруктозы, несмотря на повышенный стресс эндоплазматического ретикулума.Сахарный диабет. 2013. 62 (6): 2095–2105. DOI: 10.2337 / db12-1397. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 152. Zhang D, et al. Липогенный фактор транскрипции ChREBP опосредует индуцированную фруктозой метаболическую адаптацию для предотвращения гепатотоксичности. J Clin Invest. 2017; 127 (7): 2855–2867. DOI: 10,1172 / JCI89934. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 153. Карри DL. Влияние маннозы и фруктозы на синтез и секрецию инсулина. Поджелудочная железа. 1989. 4 (1): 2–9. DOI: 10.1097 / 00006676-1980-00002.[PubMed] [CrossRef] [Google Scholar] 154. Адамс С.Х., Стэнхоуп К.Л., Грант Р.В., Каммингс Б.П., Гавел П.Дж.. Метаболические и эндокринные профили в ответ на системную инфузию фруктозы и глюкозы у макак-резусов. Эндокринология. 2008. 149 (6): 3002–3008. DOI: 10.1210 / en.2007-1812. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 155. Блейкли С. Р., Халлфриш Дж., Райзер С., Пратер Е. С.. Долгосрочные эффекты умеренного кормления фруктозой на параметры толерантности к глюкозе у крыс. J Nutr. 1981. 111 (2): 307–314. [PubMed] [Google Scholar] 156.Бек-Нильсен Х., Педерсен О, Линдсков Х.О. Нарушение клеточного связывания инсулина и чувствительности к инсулину, вызванное кормлением с высоким содержанием фруктозы у нормальных субъектов. Am J Clin Nutr. 1980. 33 (2): 273–278. [PubMed] [Google Scholar] 157. Тер Хорст К.В., Шене М.Р., Холман Р., Ромейн Дж. А., Серли М.Дж. Влияние потребления фруктозы на чувствительность к инсулину у недиабетических субъектов: систематический обзор и метаанализ исследований диетических вмешательств. Am J Clin Nutr. 2016; 104 (6): 1562–1576. DOI: 10.3945 / ajcn.116.137786. [PubMed] [CrossRef] [Google Scholar] 158.Nagai Y, et al. Роль коактиватора гамма рецептора, активируемого пролифератором пероксисом, 1 бета (PGC-1β) в патогенезе инсулинорезистентности, индуцированной фруктозой. Cell Metab. 2009. 9 (3): 252–264. DOI: 10.1016 / j.cmet.2009.01.011. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 159. Самуэль В.Т. и др. Механизм печеночной инсулинорезистентности при неалкогольной жировой болезни печени. J Biol Chem. 2004. 279 (31): 32345–32353. DOI: 10.1074 / jbc.M313478200. [PubMed] [CrossRef] [Google Scholar] 160. Кумаширо Н. и др.Клеточный механизм инсулинорезистентности при неалкогольной жировой болезни печени. Proc Natl Acad Sci U S. A. 2011; 108 (39): 16381–16385. DOI: 10.1073 / pnas.1113359108. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 161. Фарез Р.В., младший, Зехнер Р., Ньюгард CB, Вальтер ТК. Проблема установления взаимосвязи между стеатозом печени и инсулинорезистентностью печени. Cell Metab. 2012. 15 (5): 570–573. DOI: 10.1016 / j.cmet.2012.03.004. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 163. Темплман Н.М. и др.Снижение циркулирующего инсулина повышает чувствительность к инсулину у старых мышей и увеличивает продолжительность жизни. Cell Rep. 2017; 20 (2): 451–463. DOI: 10.1016 / j.celrep.2017.06.048. [PubMed] [CrossRef] [Google Scholar] 164. Дэвис Дж. Д. Эффективность некоторых сахаров в стимулировании облизывания крыс. Physiol Behav. 1973; 11 (1): 39–45. DOI: 10.1016 / 0031-9384 (73)
-0. [PubMed] [CrossRef] [Google Scholar] 165. Авена НМ. Изучение сходных с привыканием свойств переедания с использованием модели сахарной зависимости на животных.Exp Clin Psychopharmacol. 2007. 15 (5): 481–491. DOI: 10.1037 / 1064-1297.15.5.481. [PubMed] [CrossRef] [Google Scholar] 166. Видеман СН, Надзам Г.Р., Мерфи Х.М. Последствия сахарной зависимости, абстиненции и рецидива на животной модели для здоровья человека. Nutr Neurosci. 2005. 8 (5–6): 269–276. DOI: 10.1080 / 10284150500485221. [PubMed] [CrossRef] [Google Scholar] 167. Спенглер Р., Виттковски К.М., Годдард Н.Л., Авена Н.М., Хобель Б.Г., Лейбовиц С.Ф. Опиатоподобные эффекты сахара на экспрессию генов в стимулирующих областях мозга крысы.Brain Res Mol Brain Res. 2004. 124 (2): 134–142. DOI: 10.1016 / j.molbrainres.2004.02.013. [PubMed] [CrossRef] [Google Scholar] 169. Шапиро А., Мю В., Ронкал С., Ченг К.Ю., Джонсон Р.Дж., Scarpace PJ. Резистентность к лептину, вызванная фруктозой, усиливает набор веса в ответ на последующее кормление с высоким содержанием жиров. Am J Physiol Regul Integr Comp Physiol. 2008; 295 (5): R1370 – R1375. DOI: 10.1152 / ajpregu.00195.2008. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 170. Chotiwat C, Sharp C, Teff K, Harris RBS. Питание с высоким содержанием фруктозы вызывает у крыс резистентность к лептину.Аппетит. 2007; 49 (1): 284. [Google Scholar] 171. Teff KL, et al. Пищевая фруктоза снижает циркулирующий инсулин и лептин, ослабляет постпрандиальное подавление грелина и увеличивает уровень триглицеридов у женщин. J Clin Endocrinol Metab. 2004. 89 (6): 2963–2972. DOI: 10.1210 / jc.2003-031855. [PubMed] [CrossRef] [Google Scholar] 172. Луо С., Монтероссо-младший, Сарпелле К., Пейдж КА. Дифференциальные эффекты фруктозы и глюкозы на мозг и аппетитные реакции на пищевые сигналы и решения о пищевых наградах. Proc Natl Acad Sci U S A.2015; 112 (20): 6509–6514. DOI: 10.1073 / pnas.1503358112. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 173. Page KA, et al. Влияние фруктозы и глюкозы на региональный церебральный кровоток в областях мозга, отвечающих за аппетит и пути вознаграждения. ДЖАМА. 2013. 309 (1): 63–70. DOI: 10.1001 / jama.2012.116975. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 174. Badman MK, Pissios P, Kennedy AR, Koukos G, Flier JS, Maratos-Flier E. Фактор роста фибробластов печени 21 регулируется PPARalpha и является ключевым медиатором метаболизма липидов в печени в кетотических состояниях.Cell Metab. 2007. 5 (6): 426–437. DOI: 10.1016 / j.cmet.2007.05.002. [PubMed] [CrossRef] [Google Scholar] 175. Dushay JR, Toschi E, Mitten EK, Fisher FM, Herman MA, Maratos-Flier E. Прием фруктозы резко стимулирует уровни циркулирующего FGF21 у людей. Mol Metab. 2015; 4 (1): 51–57. DOI: 10.1016 / j.molmet.2014.09.008. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 176. Инагаки Т. и др. Эндокринная регуляция реакции натощак посредством индукции фактора роста фибробластов, опосредованной PPARalpha 21.Cell Metab. 2007. 5 (6): 415–425. DOI: 10.1016 / j.cmet.2007.05.003. [PubMed] [CrossRef] [Google Scholar] 177. Санчес Дж., Палоу А., Пико С. Ответ на повторное кормление углеводами и жирами в экспрессии генов, участвующих в распределении питательных веществ и метаболизме: поразительные эффекты на индукцию фактора роста фибробластов-21. Эндокринология. 2009. 150 (12): 5341–5350. DOI: 10.1210 / en.2009-0466. [PubMed] [CrossRef] [Google Scholar] 179. Dushay J, et al. Повышенный фактор роста фибробластов 21 при ожирении и неалкогольной жировой болезни печени.Гастроэнтерология. 2010. 139 (2): 456–463. DOI: 10.1053 / j.gastro.2010.04.054. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 180. Chavez AO, Molina-Carrion M, Abdul-Ghani MA, Folli F, Defronzo RA, Tripathy D. Циркулирующий фактор роста фибробластов-21 повышен при нарушении толерантности к глюкозе и диабете 2 типа и коррелирует с мышечной и печеночной инсулинорезистентностью. Уход за диабетом. 2009. 32 (8): 1542–1546. DOI: 10.2337 / dc09-0684. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 182. Иидзука К., Такеда Дж., Хорикава Ю.Глюкоза индуцирует экспрессию мРНК FGF21 через активацию ChREBP в гепатоцитах крысы. FEBS Lett. 2009. 583 (17): 2882–2886. DOI: 10.1016 / j.febslet.2009.07.053. [PubMed] [CrossRef] [Google Scholar] 183. Фишер FM и др. Критическая роль ChREBP-опосредованной секреции FGF21 в метаболизме фруктозы в печени. Mol Metab. 2017; 6 (1): 14–21. DOI: 10.1016 / j.molmet.2016.11.008. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 184. von Holstein-Rathlou S, et al. FGF21 опосредует эндокринный контроль простого потребления сахара и предпочтения сладкого вкуса печенью.Cell Metab. 2016; 23 (2): 335–343. DOI: 10.1016 / j.cmet.2015.12.003. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 186. Танака Т. и др. Полногеномный метаанализ наблюдательных исследований показывает общие генетические варианты, связанные с потреблением макроэлементов. Am J Clin Nutr. 2013. 97 (6): 1395–1402. DOI: 10.3945 / ajcn.112.052183. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 188. Сингх А.К. и др. Гипертензия, индуцированная фруктозой: важная роль транспортеров, поглощающих хлорид и фруктозу, PAT1 и Glut5.Kidney Int. 2008. 74 (4): 438–447. DOI: 10.1038 / ки.2008.184. [PubMed] [CrossRef] [Google Scholar] 189. Джонсон Р.Дж. и др. Есть ли патогенетическая роль мочевой кислоты при гипертонии, сердечно-сосудистых и почечных заболеваниях? Гипертония. 2003. 41 (6): 1183–1190. DOI: 10.1161 / 01.HYP.0000069700.62727.C5. [PubMed] [CrossRef] [Google Scholar] 191. Джонсон Р.К. и др. Потребление диетического сахара и здоровье сердечно-сосудистой системы: научное заявление Американской кардиологической ассоциации. Тираж. 2009. 120 (11): 1011–1020. DOI: 10.1161 / CIRCULATIONAHA.109.192627. [PubMed] [CrossRef] [Google Scholar] 192. Батис Ц., Ривера Я.А., Попкин Б.М., Тайли Л.С. Оценка налога Мексики на несущественные высококалорийные продукты за первый год: обсервационное исследование. PLoS Med. 2016; 13 (7): e1002057. DOI: 10.1371 / journal.pmed.1002057. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 193. Фалбе Дж., Томпсон Х.Р., Беккер С.М., Рохас Н., Маккалок К.Э., Мадсен К.А. Влияние акцизного налога Беркли на потребление сахаросодержащих напитков. Am J Public Health.2016; 106 (10): 1865–1871. DOI: 10.2105 / AJPH.2016.303362. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]Метаболизм фруктозы — обзор
Врожденные ошибки метаболизма фруктозы
У людей было описано несколько генетических аномалий метаболизма фруктозы. Дефицит фруктокиназы приводит к высокому уровню фруктозы в крови и моче. В отсутствие фруктокиназы фруктоза может метаболизироваться до фруктозо-6-фосфата под действием гексокиназы (EC 2.7.1.1), хотя и с невысокой скоростью. Следовательно, с этой аномалией не связано никаких серьезных проблем со здоровьем. Фактически, нокаут фруктокиназы у мышей приводит к ослаблению метаболической дисрегуляции, связанной с диетами с высоким содержанием фруктозы.
Ферменты альдолазы A, B и C (EC 4.1.2.13) катализируют обратимое превращение фруктозо-1-дифосфата в глицеральдегид-3-фосфат и дигидроксиацетонфосфат, и был выявлен дефицит ферментов A и B. Альдолаза А экспрессируется в эмбриональных тканях и мышцах взрослых.Возможно, из-за важности этого фермента в гликолизе плода его дефицит приводит к умственной отсталости, низкому росту, гемолитической анемии и ненормальному внешнему виду лица. Нет известного лечения дефицита альдолазы А. Альдолаза B экспрессируется в печени, почках и кишечнике, и дефицит этого фермента более распространен и может проявляться при первом воздействии фруктозы в младенчестве или может возникнуть во взрослом возрасте. При приеме пищи, содержащей фруктозу, очевидны рвота и нарушение нормального развития.Также наблюдаются гипогликемия (в некоторых случаях тяжелая), повышение уровня мочевой кислоты в крови и нарушение функции печени. Хотя это заболевание можно эффективно лечить, исключив из рациона источники фруктозы, сахарозы и сорбита, полностью исключить эти повсеместно распространенные диетические компоненты может быть довольно сложно. Однако при отсутствии лечения может произойти серьезное повреждение органов, предположительно вызванное накоплением фруктозо-1-фосфата, который не только может быть токсичным для клеток печени и почек, но также приводит к истощению клеточного пула фосфатов.
Дефицит фруктозо-1,6-бисфосфатазы (EC 3.1.3.11) также считается генетическим нарушением метаболизма фруктозы. Этот фермент играет важную роль в ферментном комплексе, регулирующем гликолиз и глюконеогенез. У людей с дефицитом наблюдается гипогликемия, ацидоз, кетонурия, гипервентиляция и часто гипотония и гепатомегалия. Нарушается выведение с мочой многих органических кислот; в частности, уровень глицерина в моче повышен, и он полезен при диагностике этого заболевания. Лечение включает отказ от диетической фруктозы, сорбита и продолжительное голодание.
d-глицериновая ацидурия вызвана дефицитом d-глицераткиназы (EC 2.7.1.31). Было задокументировано только десять случаев с симптомами, варьирующимися от отсутствия до метаболического ацидоза, задержки развития, психомоторной отсталости, спастического тетрапареза и судорог. Отсутствие значительных симптомов у некоторых предполагает, что этот дефицит фермента по существу является доброкачественным, а дефицит других связанных ферментов может лежать в основе более серьезных симптомов.
Метаболизм фруктозы и связь с атеросклерозом, диабетом 2 типа и ожирением
Высокое потребление сахаров связано с проблемами здоровья, вызванными диетой.Содержание фруктозы в сахаре потребляется также может повлиять на здоровье, хотя степень, в которой фруктоза имеет особенно существенное негативное влияние на состоянии здоровья остается спорным. Цель этого повествовательного обзора — описать управление фруктозой в организме и обсудить роль фруктозы как фактора риска атеросклероза, диабета 2 типа и ожирения. Несмотря на некоторые положительные эффекты фруктозы, такие как высокая относительная сладость, высокий термогенный эффект и низкий гликемический индекс, высокое потребление фруктозы, особенно в сочетании с глюкозой, может в большей степени, чем аналогичное потребление глюкозы, привести к метаболическим изменениям. в печени.Повышение липогенеза de novo (DNL) и, следовательно, изменение липидного профиля крови, по-видимому, является наиболее заметным изменением. Необходимы дополнительные исследования с реалистичными уровнями потребления фруктозы, но текущая литература не указывает на то, что нормальное потребление фруктозы (приблизительно 50-60 г / день) увеличивает риск атеросклероза, диабета 2 типа или ожирения в большей степени, чем потребление других сахаров. . Однако высокое потребление фруктозы, особенно в сочетании с высоким потреблением энергии в форме глюкозы / крахмала, может иметь негативные последствия для здоровья через DNL.
1. Введение
Сахар является важным источником энергии в нашем рационе, и высокое потребление сахаров все чаще рассматривается как серьезная причина серьезных проблем со здоровьем, вызванных диетой, а именно атеросклероза, диабета 2 типа и ожирения [ 1–4]. Повышенное потребление сахаросодержащих напитков, в частности, было связано с этими проблемами со здоровьем во многих эпидемиологических исследованиях [5–7]. Основными составляющими сахаров являются фруктоза и глюкоза, которые могут присутствовать по отдельности или в комбинации, хотя чаще всего фруктоза смешивается с глюкозой в соотношении 50:50.Утверждалось, что соотношение глюкозы: фруктозы в рационе важно для некоторых эффектов сахаров [8, 9]. Таким образом, важно изучить эффекты фруктозы и глюкозы вместе, но также важно изучить эффекты одной фруктозы. Было заявлено, что фруктоза полезна, поскольку она может способствовать контролю гликемии [10, 11], но также было заявлено, что она более вредна, чем другие сахара, особенно в отношении развития атеросклероза, диабета 2 типа и ожирения [ 8].
Фруктоза содержится в различных продуктах питания. В столовом сахаре он связан с глюкозой с образованием дисахарида сахарозы, тогда как в меде он находится в моносахаридной форме. Во фруктах, ягодах и овощах фруктоза присутствует как в моносахаридной, так и в дисахаридной формах. Измеряемое как потребление калорийных подсластителей в США, потребление фруктозы было довольно стабильным на протяжении пятидесятых и шестидесятых годов, но увеличивалось с семидесятых годов до конца девяностых годов, после чего потребление снизилось [12]. Среднее потребление фруктозы в США в 2004 г. составляло 49 г в день [13].В Норвегии среднее ежедневное потребление фруктозы может быть оценено примерно в 56 г / день на основе данных исследования моделей потребления [14] и данных о составе [15], проведенного Норвежским управлением здравоохранения. Во всем мире основным источником фруктозы является сахароза, которая составляет> 90% используемых в мире энергетических подсластителей [16]. В некоторых странах, таких как США и Япония, кукурузный сироп с высоким содержанием фруктозы (HFCS) также является важным источником фруктозы. HFCS — это смесь фруктозы и глюкозы в разных концентрациях.Он может содержать до 90% фруктозы [17], но преобладающая концентрация фруктозы в коммерческих продуктах составляет 42% или 55% [9]. В США потребление HFCS резко возросло с 1970 по 1999 год, но с тех пор оно снизилось [12, 18].
Цель этой обзорной статьи — описать, как фруктоза, по сравнению с другими сахарами, регулируется организмом, тем самым разъясняя влияние фруктозы на атеросклероз, диабет 2 типа и ожирение.
2. Материалы и методы
Обзор основан на научных рецензируемых статьях, полученных с помощью несистематического поиска в базах данных PubMed и Web of Science.Первым шагом в выборе литературы было определение релевантных ключевых слов для поиска в этих базах данных. Были использованы различные комбинации следующих ключевых слов: «фруктоза», «глюкоза», «сахароза» и «сахар» в сочетании с «метаболизм», «инсулинорезистентность», «избыточный вес», «ожирение», «относительная сладость», « абсорбционная способность »,« гликемический индекс »,« липогенез de novo »,« термогенез »,« диабет 2 типа »и« регуляция аппетита ».
Следующим шагом в процессе отбора было включение или исключение статей в зависимости от их соответствия цели исследования.В ходе этого процесса первоначального отбора были включены как оригинальные, так и обзорные статьи. Списки литературы к статьям были тщательно изучены, чтобы найти какие-либо потенциально относящиеся к делу статьи, которые не были включены. Насколько это возможно, были включены только оригинальные статьи и обзоры, основанные на контролируемых испытаниях и механистических исследованиях. Если не указано иное, результаты, представленные и обсуждаемые в этой статье, являются статистически значимыми, и эксперименты проводились с контрольной группой. Хотя животные могут метаболизировать фруктозу иначе, чем люди, исследования на животных были включены, если они были признаны актуальными, например, из-за отсутствия исследований на людях.Одно возможное ограничение в этом исследовании заключается в несистематическом характере стратегии поиска, что приводит к возможности того, что соответствующие статьи не были рассмотрены.
3. Управление организмом фруктозой
3.1. Абсорбция
Хотя есть некоторая неопределенность в отношении механизмов абсорбции фруктозы, большая часть фруктозы, по-видимому, абсорбируется за счет облегченного транспорта в тощей кишке транспортером фруктозы GLUT5 [18–20]. Организм имеет ограниченную способность усваивать чистую фруктозу, поэтому потребление фруктозы может привести к нарушению всасывания [20–24].Нарушение всасывания фруктозы приводит к бактериальной ферментации, которая приводит к образованию короткоцепочечных жирных кислот (ацетат, пропионат и бутират) и газов (водород, метан и углекислый газ) [25, 26]. Эти процессы могут влиять на моторику кишечника и вызывать различные симптомы, такие как боль в животе, вздутие живота и изменение стула [27]. Наблюдались большие индивидуальные различия в абсорбционной способности фруктозы. Когда фруктоза употребляется в виде однократной пероральной дозы, максимальная абсорбционная способность составляет от 5 до 50 г [28].На эту способность влияют несколько факторов, таких как возраст и здоровье [27, 29, 30], но из диетических факторов присутствие глюкозы является наиболее важным [20, 23]. Было показано значительное увеличение всасывания фруктозы при совместном употреблении фруктозы с равным количеством глюкозы [21, 31]. По-прежнему существует неопределенность в отношении того, как глюкоза увеличивает абсорбционную способность фруктозы, но, среди прочего, это может быть связано с влиянием глюкозы на присутствие переносчиков фруктозы [20]. Фруктоза не содержится ни в каких фруктах или овощах без глюкозы [15].Эволюционно это может объяснить, почему люди не способны поглощать большие количества чистой фруктозы.
3.2. Метаболизм
После абсорбции фруктоза транспортируется по воротной вене в печень, где она эффективно всасывается клетками печени [32], в результате чего в системный кровоток попадают лишь небольшие количества. Таким образом, концентрация фруктозы в крови составляет всего около 0,01 ммоль / л, в отличие от концентрации глюкозы, которая составляет около 5,5 ммоль / л [33]. Таким образом, метаболизм фруктозы происходит в основном в печени, но фруктоза также может метаболизироваться энтероцитами.При исследовании свиней было показано, что продукция лактата кишечника из фруктозы может составлять 12% абсорбированной фруктозы [34], но функциональное значение кишечного метаболизма фруктозы у людей остается неизвестным [18]. Бьоркман и Фелиг [35] также показали, что инфузированная фруктоза (48,6 г фруктозы в жидкой форме, введенной внутривенно) метаболизируется в почках у людей, голодавших в течение 60 часов. Несмотря на искусственно завышенный уровень фруктозы в крови, это исследование по-прежнему показывает, что почки обладают относительно большой способностью метаболизировать фруктозу.Было показано, что GLUT5 экспрессируется в мембранах жировых клеток, клеток почек, мышц и мозга [32, 36], но из-за очень низкого уровня фруктозы в крови в этих тканях, вероятно, метаболизируется незначительное количество фруктозы. [9, 18].
Как обсуждалось выше, печень метаболизирует большую часть съеденной фруктозы по сравнению с примерно 15–30% проглоченной глюкозы [37, 38]. Большинство реакций фруктолиза в печени такие же, как и реакции гликолиза, но фруктоза входит в цепочку гликолитических реакций на более поздней стадии, чем глюкоза [39].Таким образом, фруктоза избегает основной контрольной стадии гликолиза, стадии фосфофруктокиназы, которая строго регулируется энергетическим статусом клетки. Первым этапом фруктолиза является фосфорилирование фруктозы фруктокиназой до фруктозо-1-фосфата. В отличие от фосфофруктокиназы при гликолизе этот фермент не ингибируется АТФ [40]. Считается, что фермент практически не регулируется, и даже при высоком энергетическом статусе печени (АТФ) фруктокиназа метаболизирует фруктозу до фруктозо-1-фосфата.Кроме того, фруктозо-1-фосфат расщепляется до дигидроксиацетонфосфата и глицеральдегида ферментом альдолазой B. Глицеральдегид может фосфорилироваться триозинкиназой с АТФ в качестве донора фосфата с образованием промежуточного гликолитического глицеральдегида 3-фосфата [41]. После этих этапов атомы углерода фруктозы следуют этапам гликолиза.
Обход стадии фосфофруктокиназы делает поток атомов углерода фруктозы через биохимические пути менее контролируемым, чем для глюкозы.Таким образом, печень будет метаболизировать фруктозу неограниченным образом, в отличие от глюкозы. Это влияет на тип и количество продуктов метаболизма, вырабатываемых печенью, и является основной причиной, по которой фруктоза и глюкоза имеют разные метаболические эффекты.
В печени фруктоза может вступать в метаболические пути: она может окисляться, превращаться в глюкозу (и гликоген), превращаться в молочную кислоту или вступать в липогенез de novo (DNL). После ночного голодания примерно 50% фруктозы, съеденной в пероральной дозе примерно 30–70 г, превращается в глюкозу в результате глюконеогенеза [42].Другие показали, что после такого же приема фруктозы окисляется около 45%; однако это включает как прямое окисление фруктозы, так и окисление глюкозы и молочной кислоты, образованной из фруктозы [43]. Другая метаболическая судьба фруктозы — это образование молочной кислоты, но это, по-видимому, происходит только при высоком потреблении фруктозы [44–48]. Употребление 72 г фруктозы — это наименьшее количество, которое, как было экспериментально продемонстрировано, приводит к образованию молочной кислоты [49]. Потребление фруктозы может также привести к образованию жирных кислот через DNL.Состав диеты может влиять на распределение фруктозы. Theytaz et al. [50] показали, что совместное употребление глюкозы с фруктозой снижает окисление и глюконеогенез атомов углерода фруктозы. Таким образом, потребление фруктозы вместе с глюкозой, по-видимому, влияет на метаболическую судьбу фруктозы. В некоторой степени этот эффект может быть связан с более высокой секрецией инсулина после приема глюкозы по сравнению с фруктозой [51]. Инсулин, среди прочего, снижает выработку глюкозы из фруктозы [52], а инсулин также стимулирует DNL [53].Степень попадания фруктозы в DNL играет ключевую роль в воздействии фруктозы на здоровье.
3.3.
De Novo Липогенез (DNL)DNL — это метаболический путь, который преобразует избыточную нежирную энергию в жир путем синтеза жирных кислот из ацетил-КоА [54]. Печень является первичным участком DNL, но DNL также может встречаться в лактирующих молочных железах и жировой ткани. Ключевые липогенные ферменты присутствуют в жировой ткани, но остается неясным, в какой степени жировая ткань влияет на общую DNL.Хотя некоторые данные указывают на то, что DNL в жировой ткани не имеет большого значения [55–57], также было показано, что она может играть более значительную роль [58], особенно после приема большого количества углеводов [59, 60]. Некоторые разногласия существуют относительно способности печени осуществлять липогенез de novo у человека [60, 61], хотя высокая способность была продемонстрирована в нескольких экспериментах [62, 63].
В результате метаболической разницы между глюкозой и фруктозой более высокий процент фруктозы по сравнению с глюкозой может быть преобразован в жир в печени посредством DNL [63].Это было показано в ряде исследований на животных и людях, в которых эти сахара потреблялись в равных количествах в аналогичных экспериментальных условиях [63–66]. Большой потенциал фруктозы в стимуляции DNL — основная причина, по которой фруктоза считается особенно вредной. Как описано ранее, печень поглощает большую часть съеденной фруктозы, а метаболизм фруктозы обходит основной контрольный этап гликолиза, что означает, что для DNL доступна большая часть фруктозы по сравнению с глюкозой.Однако нормальная диета обеспечивает в 3-5 раз больше глюкозы, чем фруктозы [12, 67], и это может повлиять на практическую значимость этого метаболического различия. Если, например, потребление глюкозы очень велико, печени может потребоваться обрабатывать большее количество глюкозы, чем фруктозы. На двух этапах фруктолиза в печени промежуточные соединения могут вступать в синтез липидов [9]. Фруктоза превращается в дигидроксиацетонфосфат, промежуточный продукт в равновесии с глицерин-3-фосфатом, который составляет основу глицерина в триглицеридах и фосфолипидах.Между тем, большая часть углеродов фруктозы метаболизируется непосредственно до пировиноградной кислоты, а затем до ацетил-КоА. При метаболизме большого количества фруктозы в ацетил-КоА количество ацетил-КоА может превышать емкость цикла лимонной кислоты митохондрий. Таким образом, высокие уровни фруктозы могут действовать как нерегулируемый источник печеночного ацетил-КоА, который является субстратом, который может проникать в DNL [9]. Кроме того, высокое потребление фруктозы, по-видимому, стимулирует экспрессию генов и активность липогенных ферментов в печени [40, 41, 66, 68].Недавно было показано, что фруктоза дает более высокое увеличение фактора роста фибробластов 21 (FGF21), чем глюкоза. FGF21 — это гормон, участвующий в гомеостазе глюкозы и липидов [69], а высокие уровни FGF21, по-видимому, связаны с метаболическими заболеваниями [70].
Очевидно, что высокое потребление фруктозы вызовет значительное увеличение активности DNL [71]. Однако мало что известно о влиянии нормального потребления фруктозы на эту активность [72]. Похоже, что не проводилось никаких исследований для оценки минимального количества или индивидуального диапазона фруктозы, которые необходимо употреблять для получения значительного увеличения DNL, что, среди прочего, может быть связано с трудностями при количественном определении DNL.Фактически, измерение DNL в печени следует считать полуколичественным [54, 73]. Чонг и др. [73] обнаружили, что 13 С-меченная фруктоза составляла лишь 0,4% триглицеридов в липопротеинах очень низкой плотности (ЛПОНП) у мужчин. Это было измерено через 6 часов после еды, содержащей 0,5 г жира / кг веса тела и 0,75 г фруктозы / кг веса тела после ночного голодания. Небольшое повышение уровня триглицеридов в крови после приема фруктозы может быть результатом замедленной секреции триглицеридов.Включение углерода из 13 C-меченой фруктозы в VLDL не всегда будет отражать DNL, потому что некоторые из вновь синтезированных жирных кислот будут задерживать секрецию [74, 75]. Уровень активности DNL после приема фруктозы, по-видимому, значительно различается у разных людей [76–78], а также в течение дня [79, 80].
Чрезмерное потребление фруктозы и, следовательно, увеличение DNL может увеличить риск заболевания, поскольку потенциально может вызывать как повышение уровня холестерина в крови, так и накопление жира в печени [81].В нескольких исследованиях на животных было продемонстрировано накопление жира в печени после высокого потребления фруктозы [82, 83], хотя большинство исследований на людях не смогли продемонстрировать такой эффект [84–86]. Фактически, Chiu et al. [87] пришли к выводу, основываясь на систематическом обзоре с метаанализом испытаний контролируемого кормления, что фруктоза не увеличивает содержание липидов в печени при изокалорийном обмене на другие углеводы. Однако они отметили, что фруктоза, потребляемая в больших дозах, может повышать содержание липидов в печени, что может быть связано с избытком энергии, а не с самой фруктозой.Браво и др. [88] провели исследование, в котором участники потребляли сахарозу или HFCS в течение десяти недель при разных уровнях потребления. В исследовании фруктоза, потребляемая как часть нормальной диеты, не способствовала увеличению содержания липидов в печени даже при высоком потреблении (уровень потребления 90-го процентиля). В исследовании, на которое ссылаются Риппе и Ангелопулос [89], не наблюдалось увеличения содержания жира в печени после потребления HFCS или сахарозы на уровне до 30% энергии в течение десяти недель. Однако Maersk et al. [90] обнаружили, что потребление 1 л подслащенного сахарозой безалкогольного напитка в день в течение шести месяцев увеличивало липиды печени у участников с избыточным весом по сравнению с потреблением того же количества молока, диетической колы и воды.Это может служить иллюстрацией того, что комбинация фруктозы и глюкозы, как в сахарозе, может привести к повышению уровня липидов в печени. Однако нельзя сделать вывод, что это эффект фруктозы, поскольку в этом исследовании не было контрольной группы глюкозы. Johnston et al. [91] не обнаружили различий в уровне триглицеридов в печени между участниками с избыточным весом, которые придерживались гиперкалорийной диеты с высоким содержанием фруктозы или высоким содержанием глюкозы. Авторы пришли к выводу, что этот результат указывает на то, что гиперкалорийное состояние, а не состав макроэлементов важны для накопления липидов в печени.Накопление жира в печени, вызванное углеводами, может привести к неалкогольной жировой болезни печени (НАЖБП) [92]. Однако до сих пор неясно, способствует ли фруктоза за счет увеличения DNL особенно благоприятной для НАЖБП [93]. Таким образом, влияние фруктозы на накопление липидов неясно, но влияние фруктозы на липидный профиль крови, по-видимому, лучше документировано.
4. Атеросклероз
Похоже, что высокое потребление фруктозы может создать неблагоприятный липидный профиль в крови через DNL [94].Основным продуктом DNL является пальмитиновая кислота [95], жирная кислота, которая, как было показано, увеличивает риск атеросклероза [96]. Жирные кислоты, образованные DNL, в основном упаковываются в VLDL, которые доставляются в кровоток. Это, в свою очередь, может повысить уровень липопротеинов низкой плотности (ЛПНП) в крови. В нескольких исследованиях фруктоза в большей степени, чем глюкоза, увеличивала уровни триглицеридов в крови [51, 65, 97, 98] и ЛПНП [65, 99-103]. Aeberli et al. [104] показали, что фруктоза увеличивает небольшие плотные ЛПНП, тип ЛПНП, который, в частности, может быть связан с сердечно-сосудистым риском [105].Уровень липопротеинов высокой плотности (ЛПВП) в крови, по-видимому, не зависит от фруктозы [100, 101]. В большинстве исследований потребление фруктозы> 100 г / день было необходимо для наблюдения неблагоприятного воздействия на липидный профиль [51, 65, 85, 98, 100, 106–108]. Однако в недавнем исследовании Aeberli et al. [109], ежедневное потребление около 77 г фруктозы и 34 г глюкозы в течение 3 недель привело к повышению уровня общего холестерина и ЛПНП в крови здоровых молодых мужчин по сравнению с ежедневным потреблением примерно 109 г глюкозы и 28 г фруктозы. за тот же период времени.Тот факт, что обе группы также потребляли неизвестное количество крахмала, и тот факт, что потребление пищи не контролировалось, снижают достоверность этих результатов. Maersk et al. [90] обнаружили, что потребление 50 г фруктозы в день вместе с 50 г глюкозы может отрицательно влиять на уровень триглицеридов в крови. Отсутствие глюкозы в контрольной группе затрудняет вывод о том, что это эффект фруктозы. Напротив, Lowndes et al. [110] не обнаружили отрицательного воздействия на липидный профиль у лиц с избыточным весом или ожирением, потребляющих HFCS или сахарозу, включенную в эукалорийную диету в течение десяти недель на уровнях, соответствующих 25-м и 50-м процентилям потребления фруктозы взрослыми.Используя современные знания, не представляется, что потребление только умеренных количеств фруктозы (<50 г / день) приведет к неблагоприятному липидному профилю крови [86, 111].
Из-за незначительных уровней фруктозы в периферической крови, как описано выше, только глюкоза потенциально может быть субстратом для DNL в жировой ткани. Хотя DNL в жировой ткани кажется небольшим, как обсуждалось ранее, глюкоза из-за своего присутствия в крови и повышения уровня инсулина в крови, вероятно, в большей степени, чем фруктоза, будет стимулировать DNL в жировой ткани.Потребление глюкозы в количествах, превышающих общую способность хранения гликогена и окисления глюкозы, может, таким образом, увеличить DNL в жировой ткани больше, чем такое же количество фруктозы. Хотя жир, образующийся в печени, должен переноситься в кровь в виде липопротеинов, этого можно избежать, если жир образуется непосредственно в жировой ткани. Принимая во внимание известные негативные воздействия на здоровье остатков липопротеинов, наличие DNL в жировой ткани может быть предпочтительнее, чем DNL в печени. Это может свидетельствовать о различии метаболизма между глюкозой и фруктозой при потреблении большого количества сахаров.
Еще одно возможное различие между фруктозой и глюкозой в факторах риска атеросклероза — это влияние этих сахаров на уровень мочевой кислоты в крови. Повышенный уровень мочевой кислоты был связан с атеросклерозом в эпидемиологических исследованиях, но причинно-следственная связь неясна [112–114]. Фруктоза, по-видимому, увеличивает уровень мочевой кислоты в крови в большей степени, чем глюкоза, особенно при большом потреблении и при потреблении в качестве избыточной энергии [86, 115, 116]. Потребление 0,5 г фруктозы / кг массы тела — это наименьшее количество, которое, как показано, приводит к образованию мочевой кислоты [117].Повышенный уровень мочевой кислоты в крови теоретически может привести к повышению артериального давления, поскольку мочевая кислота ингибирует фермент в эндотелиальных клетках артерий, называемый эндотелиальной синтазой оксида азота (eNOS). Активированный eNOS приводит к увеличению выработки оксида азота (NO), важного вазодилататора. Таким образом, ингибирование eNOS может привести к сужению сосудов. Хотя было показано, что только 0,5 г фруктозы / кг массы тела вызывает образование мочевой кислоты, а повышенный уровень мочевой кислоты теоретически может повышать артериальное давление, результаты исследований влияния фруктозы на артериальное давление очень противоречивы [65, 72, 118, 119 ].Среднее потребление фруктозы, похоже, не приводит к повышению артериального давления [111, 120, 121]. Отсутствие причинной связи между уровнем мочевой кислоты и атеросклерозом затрудняет выводы о влиянии фруктозы.
5. Диабет 2 типа
Высокое потребление сахаросодержащих напитков, содержащих фруктозу в качестве одного из основных типов моносахаридов, было связано с развитием диабета 2 типа [5, 122]. Хотя эта связь не доказывает причинно-следственную связь, важно изучить роль фруктозы в развитии диабета 2 типа.Центральное место в понимании диабета 2 типа занимает влияние питательных веществ на гомеостаз глюкозы в крови. Фруктоза должна превращаться в глюкозу в печени, чтобы вызвать повышение уровня глюкозы в крови. Поскольку преобразование требует времени и только часть фруктозы образует глюкозу, фруктоза увеличивает уровень глюкозы в крови меньше, чем аналогичный уровень глюкозы [51]. Таким образом, гликемический индекс фруктозы составляет всего 23 [10]. Это, вместе с отсутствием стимуляции клеток поджелудочной железы β [123], дает более низкую секрецию инсулина после приема фруктозы по сравнению с глюкозой [51, 124, 125].Эти эффекты положительны, потому что они способствуют гомеостазу глюкозы в крови. Кроме того, было показано, что умеренное количество фруктозы положительно влияет на гликемический контроль [86, 126, 127]. Однако утверждается, что фруктоза также может отрицательно влиять на гомеостаз глюкозы в крови, вызывая инсулинорезистентность в печени [9]. Имеются данные о том, что высокое потребление фруктозы может вызывать инсулинорезистентность у животных [128, 129], но несколько исследований на людях не смогли продемонстрировать такую связь [103, 130–132].Оперативное определение инсулинорезистентности или чувствительности кажется неясным, и для ее измерения использовалось множество различных методов [133]. Таким образом, сложно сравнивать исследования инсулинорезистентности. В исследованиях на людях, в которых фруктоза, как сообщалось, вызывает инсулинорезистентность, суточное потребление фруктозы составляло 110 г [109], примерно 250 г [134], 80 г [135] и 138 г [65]. . Это может указывать на то, что потребление фруктозы должно быть высоким, чтобы потенциально вызвать инсулинорезистентность [86].В исследованиях Aeberli et al. [109], Stanhope et al. [65] и Beck-Nielsen et al. [134], общее потребление пищи не контролировалось. Таким образом, наблюдаемый эффект фруктозы также мог быть вызван различиями в потреблении пищи между контрольной и экспериментальной группами. Во всех исследованиях, в которых была показана инсулинорезистентность, фруктозу употребляли вместе с глюкозой или крахмалом, поэтому наблюдения также могли быть результатом комбинации фруктозы и глюкозы. Был предложен ряд гипотез о том, как фруктоза может вызывать инсулинорезистентность в печени.Накопление липидов в печени [136–138], мета воспалительные процессы [83] и окислительный стресс [139] являются возможными механизмами индуцированной фруктозой резистентности к инсулину посредством ингибирующего фосфорилирования рецептора инсулина или сигнальных молекул, участвующих в передаче сигналов инсулина [ 9]. Однако исследований на людях слишком мало, и они слишком расходятся, чтобы можно было сделать однозначный вывод о связи между потреблением фруктозы и инсулинорезистентностью. Необходимы более долгосрочные исследования, в которых ежедневное потребление фруктозы является умеренным.
6. Ожирение
Спорный вопрос, является ли фруктоза менее насыщающей, чем другие сахара, и, таким образом, может способствовать ожирению из-за большого количества потребляемой пищи. В исследовании Page et al. [92], магнитно-резонансные (МР) изображения были сделаны человеческого мозга после употребления 75 г фруктозы или напитка из глюкозы. Глюкоза, но не фруктоза, снижает активность (регионарный церебральный кровоток) в гипоталамусе в областях, участвующих в регуляции энергии и системах вознаграждения; это, вероятно, показатель насыщения и может указывать на то, что фруктоза насыщает меньше, чем глюкоза.Фруктоза также в меньшей степени, чем глюкоза, увеличивает уровни инсулина [51], лептина [51, 140], желудочного ингибитора полипептида [141] и глюкагоноподобного пептида-1 [92] в крови, в то же время снижают уровень грелина в меньшей степени [51]. Хотя эти гормональные эффекты могут указывать на то, что фруктоза менее насыщает, чем глюкоза, это не было подтверждено в исследованиях способности фруктозы насыщать. В таких исследованиях было показано, что фруктоза в большей степени снижает аппетит, чем глюкоза, когда потребление происходит перед едой [142, 143], или что нет разницы во влиянии фруктозы и глюкозы на аппетит [144, 145].Таким образом, влияние фруктозы на аппетит остается неясным.
Хотя вполне возможно, что фруктоза из-за отсутствия стимуляции сигналов сытости может способствовать ожирению, фруктоза обладает несколькими свойствами, которые действуют против ожирения. Как упоминалось ранее, тонкий кишечник имеет ограниченную способность абсорбировать фруктозу. Это может привести к мальабсорбции, по крайней мере, если потребляются большие количества и потребление происходит без питательных веществ, обеспечивающих глюкозу. Нарушение всасывания фруктозы приведет к тому, что меньше фруктозы попадет в кровоток, и, следовательно, меньше энергии будет доступно клеткам.Таким образом, нарушение всасывания будет действовать против ожирения. В многочисленных исследованиях также было показано, что фруктоза обладает большим термогенным эффектом, чем глюкоза [46, 146–148]. Это означает, что после употребления фруктозы организм использует больше энергии, чем глюкозы, поэтому меньше энергии будет доступно для хранения в виде жира. Относительная сладость фруктозы также больше, чем у глюкозы и сахарозы [149, 150]. Хотя он будет уменьшаться с повышением температуры [151, 152], высокая относительная сладость позволяет использовать меньшее количество фруктозы, чем глюкоза и сахароза, для достижения особой сладости в большинстве случаев.Судя по этим свойствам, нельзя сказать, что фруктоза дает больший жир, чем другие сахара. Это также согласуется с экспериментальными исследованиями взаимосвязи между потреблением фруктозы и ожирением у животных [153, 154] и людей [65, 86, 106, 155].
7. Окисление субстрата
Было высказано предположение, что фруктоза может ингибировать окисление липидов [146]. Для окисления липидов в печени это логично, потому что печень получает энергию от фруктозы и, следовательно, не нуждается в окислении жира.Фруктоза также может увеличивать DNL. Было бы нецелесообразно одновременно образовывать и расщеплять жир в печени, а повышенный уровень малонил-КоА (из-за активной DNL) будет ингибировать β -окисление [41, 156]. Было бы также логично, если бы фруктоза увеличивала общее окисление липидов в организме больше, чем глюкоза, из-за меньшего вклада фруктозы как внепеченочного источника энергии. Однако в некоторых исследованиях было показано, что фруктоза увеличивает коэффициент дыхания (RQ), соотношение выдыхаемого CO 2 к потребляемому O 2 , [157] больше, чем глюкоза.Это указывает на то, что фруктоза в большей степени, чем глюкоза, снижает общее окисление липидов в организме и увеличивает общее окисление углеводов. Blaak и Saris [158] провели исследование, в котором участники съели 75 г фруктозы, крахмала или глюкозы после 12-часового голодания в перекрестном исследовании. Фруктоза привела к значительно большему увеличению RQ, измеренному через 6 часов после приема внутрь, чем глюкоза и крахмал. Tappy et al. [146] также показали большее увеличение RQ через 4 часа после приема 75 г фруктозы по сравнению с такими же здоровыми участниками, которые потребляли 75 г глюкозы.Schwarz et al. [46] провели аналогичное исследование (75 г фруктозы / глюкозы) с тем же результатом. Эти результаты можно объяснить тем фактом, что фруктоза больше, чем глюкоза, попадает в DNL при определенных условиях. В связи с тем, что RQ варьируется для разных субстратов (например, 1 для углеводов и 0,7 для жиров), RQ используется для определения источника окисления субстрата [159]. Однако во время активного DNL CO 2 производится без использования O 2 . Одновременное возникновение DNL и окисления углеводов может привести к значениям RQ больше 1 [159].Поэтому повышенный RQ, вызванный DNL, может быть неверно истолкован как пониженное окисление липидов и повышенное окисление углеводов. Такое неверное толкование могло иметь место в исследованиях, описанных выше. Таким образом, влияние фруктозы на общее окисление субстратов в организме остается неясным.
8. Обсуждение
Распределение фруктозы по метаболическим путям, особенно DNL, имеет ключевое значение для воздействия фруктозы на здоровье. Распределение зависит от количества потребляемой фруктозы, продолжительности воздействия фруктозы, состава диеты / приема пищи и от того, проводилось ли измерение после приема пищи, после поглощения или в условиях голодания.Также важны индивидуальные физиологические, ферментативные и эндокринные факторы. Состав диеты и количество съеденной и усвоенной фруктозы будут в центре внимания этого обсуждения.
Нарушение всасывания фруктозы влияет на количество абсорбированной фруктозы и, таким образом, может быть важным смешивающим фактором в исследованиях, в которых факторы, влияющие на абсорбционную способность, не принимались во внимание [39]. Truswell et al. [21] показали, что потребление 50 г фруктозы приводило к нарушению всасывания более чем у 50% участников исследования.Таким образом, результаты исследований мальабсорбции фруктозы могут быть неточными из-за индивидуальных различий в абсорбционной способности фруктозы. Поскольку тонкий кишечник обладает большой абсорбционной способностью для глюкозы и ограниченной для фруктозы, сложно сравнивать фруктозу с глюкозой как единственным источником углеводов. В будущих исследованиях это следует контролировать, например, с помощью водородного дыхательного теста для оценки мальабсорбции фруктозы.
Состав рациона и особенно количество глюкозы / крахмала могут влиять на воздействие фруктозы на здоровье.Поскольку фруктоза присутствует вместе с глюкозой в большинстве пищевых продуктов, более практично и актуально рассматривать эффекты фруктозы и глюкозы вместе, чем эффекты одной фруктозы. Было показано большее увеличение DNL после одновременного употребления фруктозы и глюкозы (50:50 глюкоза: фруктоза), а не такого же количества, как у чистой глюкозы [63]. Употребление глюкозы с фруктозой, вероятно, повлияет на воздействие фруктозы на здоровье, стимулируя приток фруктозы к DNL [50]. Этот эффект может быть связан как с повышенной абсорбционной способностью фруктозы при совместном употреблении с глюкозой, и, следовательно, с большей доступностью атомов углерода фруктозы, идущих в направлении DNL, так и с повышенным уровнем инсулина в крови, когда глюкоза присутствует в рационе.Инсулин прямо или косвенно стимулирует DNL, подавляя другие важные метаболические пути фруктозы, такие как глюконеогенез. Также вероятно, что глюкоза будет конкурировать с фруктозой в качестве источника энергии для энтероцитов и печеночных клеток, тем самым делая больше фруктозы доступной для DNL. Было показано, что совместное употребление глюкозы с фруктозой снижает окисление и глюконеогенез атомов углерода фруктозы [50]. Таким образом, кажется, что комбинация фруктозы и глюкозы особенно неудачна, хотя фруктоза не является предпосылкой для DNL.Неясно, какое количество глюкозы / крахмала должно быть включено во фруктозу, чтобы этот эффект был значительным.
Количество потребляемой фруктозы имеет большое значение для воздействия фруктозы на здоровье. Негативное воздействие фруктозы на здоровье в основном было продемонстрировано при высоком потреблении, и несколько исследований показали, что среднее потребление фруктозы не вызывает проблем со здоровьем [12, 110, 111, 160]. Хотя среднесуточное потребление фруктозы составляет 50–60 г / день, некоторые люди будут потреблять большие количества [161, 162], поэтому негативные последствия фруктозы для здоровья могут иметь значение, по крайней мере, для части населения.Многие из текущих исследований плохо подходят для определения рисков для здоровья от фруктозы, потому что (а) потребление фруктозы нереально высокое, (б) фруктоза вводится изолированно, а не в смеси с другими углеводами, как на практике, и / или (в) исследования проводятся на животных. Различия между физиологией человека и животных ограничивают их применимость к людям. Существует потребность в дополнительных исследованиях на людях в условиях, более похожих на то, как обычно потребляется фруктоза. Такие исследования, особенно связанные с DNL, будут необходимы, чтобы понять эффекты нормального потребления фруктозы.Влияние фруктозы на уровни триглицеридов и холестерина в крови, накопление жира в печени и передачу сигналов инсулина, по-видимому, связано со степенью проникновения фруктозы в DNL. Удивительно, но удовлетворительных исследований для оценки доли фруктозы, попадающей в DNL, при разных уровнях потребления не проводилось. Такие исследования должны быть проведены и должны включать потребление как чистой фруктозы, так и фруктозы вместе с глюкозой.
Также важно отметить, что, несмотря на различия в метаболизме глюкозы и фруктозы, потребление глюкозы намного превышает потребление фруктозы в рационе человека [12].Этот количественный аспект необходимо учитывать при сравнении воздействия глюкозы и фруктозы на здоровье.
9. Заключение
Несмотря на то, что опубликованной литературы о физиологических эффектах фруктозы у людей мало, текущая литература не указывает на то, что нормальное потребление фруктозы (приблизительно 50-60 г / день) увеличивает риск атеросклероза, типа 2 диабет или ожирение больше, чем потребление других сахаров. Однако высокое потребление фруктозы, особенно в сочетании с высоким потреблением энергии в форме глюкозы / крахмала, может иметь негативные последствия для здоровья через DNL.Совершенно очевидно, что необходимы дополнительные исследования, особенно исследования при более реалистичных уровнях потребления фруктозы.
Конфликт интересов
Авторы заявляют об отсутствии конфликта интересов в отношении публикации данной статьи.
Насколько вредна фруктоза? | Американский журнал клинического питания
В этом выпуске журнала есть еще одна тревожная статья о биологии фруктозы (1). Почему вызывает беспокойство фруктоза? Во-первых, он слаще глюкозы или сахарозы.Во фруктах он служит маркером для продуктов, богатых питательными веществами. Однако в безалкогольных напитках и других «сладостях» фруктоза служит поощрением сладкого вкуса, который обеспечивает «калорийность», зачастую без каких-либо дополнительных питательных свойств. Во-вторых, потребление безалкогольных напитков, содержащих кукурузный сироп с высоким содержанием фруктозы (HFCS) или сахарозу, выросло параллельно с эпидемией ожирения, что предполагает связь (2). В-третьих, статья в этом выпуске журнала (1) и другая статья, опубликованная в другом месте в прошлом году (3), указывают на то, что диетическая фруктоза является потенциальным фактором риска сердечно-сосудистых заболеваний.
Потребление фруктозы с пищей значительно увеличилось с 1970 по 2000 год. За этот период количество доступных «добавленных сахаров» увеличилось на 25% (4). Непрерывное исследование потребления пищи отдельными лицами с 1994 по 1996 год показало, что средний человек ежедневно потреблял 79 г добавленных сахаров (что эквивалентно 316 ккал / день или 15% калорийности), примерно половину из которых составляла фруктоза. Что еще более важно, люди, входящие в верхнюю треть потребителей фруктозы, потребляют 137 г добавленных сахаров в день, а те, кто входит в верхние 10%, потребляют 178 г / день, причем половину этого количества составляет фруктоза.Если есть проблемы со здоровьем, связанные с фруктозой, то повышенное потребление может усугубить эти проблемы.
До того, как европейцы столкнулись с Новым Светом 500 лет назад и началась мировая сахарная промышленность, фруктоза в рационе человека была ограничена несколькими продуктами. Например, мед, финики, изюм, патока и инжир содержат более 10% этого сахара, тогда как содержание фруктозы 5–10% по весу содержится в винограде, сырых яблоках, яблочном соке, хурме и чернике. . Молоко, основная пища для младенцев, по сути, не содержит фруктозы, как и большинство овощей и мяса, что указывает на то, что до массового производства сахара люди мало контактировали с фруктозой.
Большая часть фруктозы в американском рационе поступает не из свежих фруктов, а из HFCS или сахарозы (сахара), которые содержатся в безалкогольных напитках и сладостях, которые обычно содержат мало других питательных веществ (2). Потребление безалкогольных напитков, которые обеспечивают большую часть этой фруктозы, резко возросло за последние 6 десятилетий, увеличившись с 90 порций на человека в год (≈2 порции в неделю) в 1942 году до 600 порций в год (≈ 2 порции в день) в 2000 г. (5). Более 50% детей дошкольного возраста употребляют некоторые подслащенные калорийными напитками (6).Дети этого возраста обычно не подвергаются воздействию фруктозы, не говоря уже о таких больших количествах. Поскольку и HFCS, и сахароза являются «средствами доставки фруктозы», нагрузка фруктозы увеличивалась параллельно с использованием сахара.
Фруктоза является промежуточным звеном в метаболизме глюкозы, но биологической потребности во фруктозе нет. При проглатывании сама по себе фруктоза плохо всасывается из желудочно-кишечного тракта и почти полностью выводится печенью — циркулирующая концентрация составляет ≈0.01 ммоль / л в периферической крови по сравнению с 5,5 ммоль / л для глюкозы.
Фруктоза несколько отличается от глюкозы, другой половины молекулы сахарозы (сахара) (4). Фруктоза всасывается из желудочно-кишечного тракта по другому механизму, чем глюкоза. Глюкоза стимулирует высвобождение инсулина из изолированной поджелудочной железы, а фруктоза — нет. Большинство клеток имеют лишь небольшое количество переносчика глют-5, который переносит фруктозу в клетки. Фруктоза не может проникнуть в большинство клеток, потому что им не хватает глют-5, тогда как глюкоза транспортируется в клетки глют-4, инсулинозависимой транспортной системой.Наконец, попав в клетку печени, фруктоза может попасть в пути, которые обеспечивают глицерин, основу триацилглицерина. Растущее количество фруктозы, получаемой из сахарозы или HFCS, вызывает вопросы о том, как дети и взрослые реагируют только на фруктозу или когда она сопровождается глюкозой. В одном исследовании потребление пищи с высоким содержанием фруктозы снижало 24-часовые концентрации инсулина и лептина в плазме и повышало уровень триацилглицеринов после приема пищи натощак у женщин, но не подавляло циркуляцию грелина, главного гормона, стимулирующего аппетит (4).
Фруктоза метаболизируется, в первую очередь, в печени, путем фосфорилирования в положении 1, процесса, который обходит стадию фосфофруктокиназы, ограничивающую скорость (4). Таким образом, метаболизм фруктозы в печени способствует липогенезу, и неудивительно, что в нескольких исследованиях были обнаружены изменения в циркулирующих липидах, когда субъекты едят пищу с высоким содержанием фруктозы (4). В исследовании, проведенном Aeberli et al (1), диетические факторы, особенно фруктоза, изучались в зависимости от индекса массы тела, соотношения талии и бедер, липидного профиля плазмы и размера частиц ЛПНП у 74 швейцарских школьников в возрасте от 6 до 6 лет. 14 лет.В этом исследовании триацилглицерины в плазме были выше, концентрации холестерина ЛПВП были ниже, а размер частиц липопротеинов (ЛПНП) был меньше у детей с избыточным весом, чем у детей с нормальным весом. У более полных детей размер частиц ЛПНП был меньше, и даже после контроля ожирения потребление фруктозы с пищей было единственным диетическим фактором, связанным с размером частиц ЛПНП. В этом исследовании именно свободная фруктоза, а не сахароза, была связана с влиянием размера частиц ЛПНП. Исследования на грызунах, собаках и нечеловеческих приматах, употребляющих пищу с высоким содержанием фруктозы или сахарозы, неизменно показывают гиперлипидемию (4).Текущий отчет Aeberli et al. Предполагает, что более высокое потребление фруктозы детьми школьного возраста может иметь пагубные последствия для их будущего риска сердечно-сосудистых заболеваний за счет уменьшения размера частиц ЛПНП. Интересно, что это исследование не обнаружило связи диетической фруктозы с триацилглицеринами, но обнаружило связь с более липидными частицами, холестерином ЛПНП. В другом недавнем отчете была предложена гипотеза, связывающая потребление фруктозы с давно известной взаимосвязью между мочевой кислотой и сердечными заболеваниями (3).АДФ, образованный из АТФ после фосфорилирования фруктозы в положении 1, может далее метаболизироваться до мочевой кислоты. Метаболизм фруктозы в печени стимулирует выработку мочевой кислоты, которая использует оксид азота, ключевой модулятор функции сосудов (3). Исследования Aeberli et al и Nakagawa et al предполагают, что связь фруктозы со здоровьем требует переоценки.
У автора не было личного или финансового конфликта интересов.
ССЫЛКИ
1Aeberli
I
,Zimmermann
MB
,Molinari
L
и др..Потребление фруктозы является предиктором размера частиц ЛПНП у школьников с избыточным весом
.Am J Clin Nutr
2007
;86
:1174
—8
,2Брей
GA
,Nielsen
SJ
,Popkin
BM
.Потребление кукурузного сиропа с высоким содержанием фруктозы в напитках может сыграть роль в эпидемии ожирения
.Am J Clin Nutr
2004
;79
:537
—43
.3Накагава
T
,Hu
H
,Жариков
S
и др. .Причинная роль мочевой кислоты в метаболическом синдроме, индуцированном фруктозой
.Am J Physiol (почечная физиология)
2006
;290
:F625
—31
.4Гавел
PJ
.Диетическая фруктоза: последствия для нарушения регуляции энергетического гомеостаза и липидного / углеводного обмена
.Nutr Ред.
2005
;63
:133
—57
,5Вартаниан
LR
,Schwartz
MB
,Brownell
KD
.Влияние потребления безалкогольных напитков на питание и здоровье: систематический обзор и метаанализ
.Am J Public Health
2007
;97
:667
—75
,6Harnack
L
,Stang
J
,Story
M
.Потребление безалкогольных напитков детьми и подростками в США: последствия для питания
.J Am Diet Assoc
1999
;99
:436
—41
.© 2007 Американское общество клинического питания
|
Фруктоза и ее влияние на наш организм
Кукурузный сироп с высоким содержанием фруктозы не является безобидным химическим веществом.
В то время как фруктоза кажется лучшей альтернативой диетической глюкозе из-за ее инсулино-сберегающих свойств, начинают появляться исследования, связывающие потребление HFCS с множеством проблем, связывая их с стремительно растущим числом: дегенеративных заболеваний, метаболического синдрома. , ожирение, повышение уровня триглицеридов (дислипидемия), высокое кровяное давление и диабет 2 типа. Кроме того, HFCS участвует в нарушении регуляции иммунной системы с чрезмерной экспрессией воспаления в организме (окислительный стресс и воспалительные цитокины).
Другие исследования также связали нарушение пищеварения фруктозы с повышением уровня амилазы в сыворотке (пищеварительный фермент, секретируемый поджелудочной железой), снижением уровня фолиевой кислоты в сыворотке (фолат является одним из витаминов группы B. Он жизненно важен для правильного роста нервной системы у развивающегося плода). и металлическая депрессия, особенно у женщин.
Фруктоза — ценное и необходимое питательное вещество. Проблема не в фруктозе, а в ее чрезмерно обильном количестве в пищевых продуктах, производимых нами. Фруктоза, содержащаяся во фруктах и овощах, медленно всасывается в кровоток, ее абсорбция затрудняется растворимой клетчаткой, естественным образом содержащейся в этих продуктах.Напротив, HFCS и кристаллическая фруктоза нарушают метаболизм в печени, который, наряду с избытком глюкозы, резко увеличивает уровень сахара в крови и истощает нашу поджелудочную железу. Это подавляющее количество фруктозы и последующий всплеск уровня сахара в крови, которые подрывают метаболизм и иммунную систему нашего организма, чего мы не до конца понимаем.
Наши современные диеты оказались одним большим неконтролируемым экспериментом над здоровьем нашей нации. Промышленный сахар — не друг нашему телу. Чтобы узнать больше об опасностях искусственного сахара, мы рекомендуем книгу Нэнси Эпплтон, доктор философии (питание), под названием Suicide by Sugar.И для дальнейшего чтения из очень авторитетного источника.
Как фруктоза всасывается в пищеварительном тракте и как она метаболизируется в организме?
Фруктоза — это простой сахар или так называемый моносахарид. Он содержится в 3 основных формах: во фруктах и некоторых овощах, как компонент сахарозы (дисахарид, состоящий из глюкозы + фруктозы) и как природный полимер фруктозы (цепочка молекул фруктозы), называемый фруктаном (содержится в пшенице) и немного овощей).
В то время как глюкоза легко проходит через барьер кишечной мембраны, фруктозе требуется помощь транспортной системы — основной из них называется GLUT5.
Всасывание фруктозы через кишечную мембрану дополнительно облегчается глюкозой. Когда глюкоза и фруктоза присутствуют в кишечнике в соотношении один к одному, всасывание фруктозы происходит с максимальной эффективностью. Сахароза (столовый сахар), химически состоящая из равных количеств глюкозы и фруктозы, обычно полностью всасывается в тонком кишечнике.
Однако в случае таких продуктов, как кристаллическая фруктоза или HFCS, два фактора создают проблемы с абсорбцией фруктозы. Во-первых, низкое соотношение глюкозы и фруктозы в этих продуктах означает, что значительная часть съеденной фруктозы останется неабсорбированной и продолжит свое путешествие по пищеварительному тракту. Во-вторых, GLUT5 быстро забивается, когда он представлен большим количеством фруктозы, тем самым ограничивая скорость прохождения фруктозы в кровоток. Как вы узнаете, неабсорбированная фруктоза может стать взрывоопасной проблемой.
Верхняя часть тонкой кишки имеет наибольшую концентрацию GLUT5 и, следовательно, является местом большей части всасывания фруктозы. Количество единиц GLUT5 варьируется от человека к человеку в зависимости от генетической наследственности — факт, который объясняет различия между людьми в их способности усваивать одинаковое количество фруктозы.
И независимо от генетического распределения GLUT5, каждый человек может в определенной степени активировать свой GLUT5 — то есть, когда человек ест больше фруктозы, организм имеет способность компенсировать это за счет производства большего количества GLUT5 — или, наоборот, когда человек сокращает его потребление. при потреблении фруктозы организм избавляется от лишних ненужных единиц GLUT5.Думайте об этом как о бодибилдере, который может увеличить мышечную массу, поднимая тяжелые веса — он может быть не самым сильным парнем (это зависит от генетики), но он может добавить мышечную массу своему телу и, наоборот, потеряет лишние мышцы после ухода из обычного спортзала тренировки.
Следует сделать важное замечание: недавние научные данные показали, что у людей с диабетом 2 типа наблюдается значительная повышенная регуляция экспрессии GLUT5, что дает их кишечнику повышенную способность абсорбировать моносахараиды (глюкозу и фруктозу).Это подчеркивает важность диетических модификаций для борьбы с диабетом.
Другая опубликованная статья обнаружила гендерные различия в способности усваивать фруктозу. Было обнаружено, что женщины чаще страдают нарушением пищеварения фруктозы, чем мужчины.
В то время как глюкоза может быть использована (метаболизирована) практически каждой клеткой человеческого тела, фруктоза — нет. Фруктозу необходимо обрабатывать и хранить в печени в качестве резервного источника энергии, называемого гликогеном. Как только вместимость печени заполняется, избыток фруктозы превращается печенью в различные продукты; один из основных продуктов — триглицериды.Триглицериды далее превращаются печенью в липопротеины очень низкой плотности (ЛПОНП), которые высвобождаются для хранения в жировых клетках и мышцах.
Переработка избытка фруктозы печенью не является безэнергетической задачей. За усложнение работы печени приходится расплачиваться за последствия, одним из которых является незначительный сдвиг в кислотно-щелочном балансе в организме, что приводит к переходу в кислую среду. Диета с низким содержанием фруктов и овощей также усугубляет проблему. Чистый кислотный сдвиг в химии нашего тела может иметь многочисленные последствия, которые негативно сказываются на нашем здоровье — одно конкретное последствие связано со здоровьем костей и развитием остеопороза.
Для получения дополнительной информации.
Избыток фруктозы в кишечнике: каковы последствия?
Как видите, скорость всасывания фруктозы ограничена количеством единиц GLUT5, с которыми мы родились, и / или количеством потребляемой глюкозы (или, скорее, соотношением глюкозы к фруктозе).
Мальабсорбция фруктозы зависит от дозы (и глюкозы) — это означает, что вам, возможно, удастся избежать употребления небольшого количества фруктозы, но по мере увеличения концентрации фруктозы, вы будете усиливать мальабсорбцию.
Кроме того, многие промышленные продукты могут содержать большое количество фруктозы и мало или совсем не содержать глюкозы. Когда молекула глюкозы не присутствует одновременно с молекулой фруктозы, мало или совсем немного фруктозы будет абсорбироваться и, следовательно, будет проходить в толстую кишку.
Наконец, проблема заключается в большом количестве потребляемой фруктозы. Помните, среднестатистический американец съедает 80 граммов фруктозы каждый день — все это не может попасть из кишечника в кровоток (и, возможно, это хорошо для нашей печени!).Затем весь этот избыток фруктозы продолжает свой путь и попадает в пищеварительный тракт.
Есть и другие организмы, живущие внутри нас, которые рады найти дополнительную фруктозу — бактерии в нашем толстом кишечнике! В нашем кишечнике обитают миллиарды бактерий. Половина нашего стула состоит из выброшенных бактерий. Кроме того, существует около 400 видов кишечных бактерий — каждый из нас несет уникальное соотношение этих бактерий. Предпочтительным питательным веществом для кишечных бактерий являются молекулы сахара.
Когда фруктоза встречается с бактериями, начинается пир! Бактерии быстро сбраживают фруктозу в различных пропорциях различных газов, таких как водород, метан, диоксид углерода, сероводород и короткоцепочечные жирные кислоты. Каждый вид бактерий выделяет один из этих газов и, в зависимости от того, каких видов слишком много в толстой кишке, определяется характер вашего избыточного газа. Большинство людей в основном производят водород, тогда как меньшее количество людей в основном выделяет метан. Ферментирующие газы быстро накапливаются, вызывая вздутие живота, вздутие живота, дискомфорт в кишечнике или боль в животе.
Избыток фруктозы, попадающий в кишечник, не только питает бактерии, но и притягивает с собой лишнюю воду (осмотический эффект), которая оказывает слабительное действие на кишечник, вызывая диарею. К этому добавляется еще один побочный продукт метаболизма кишечных бактерий — короткоцепочечные жирные кислоты, которые также служат для привлечения большего количества воды в кишечник.
Метан не инертен. Генри Линь и Марк Пиментел в больнице Седарс Синай в Лос-Анджелесе провели ряд экспериментов, которые показали, что избыток метана в толстой кишке является биологически активным и способен нарушать двигательную активность (моторику) толстой кишки, вызывая спазмы и спазмы. спазмы кишечника и замедление прохождения стула — все это приводит к боли в животе и запору.
Возникает соблазн предположить, что причина, по которой некоторые люди могут справляться с избытком фруктозы в рационе, не страдая от ее пагубного воздействия на кишечник, может быть связана с природой их бактерий толстой кишки. Их преобладающий урожай насекомых может оказаться защитным, обеспечивая меньшую нагрузку на нижние отделы пищеварительного тракта — независимо от неправильного питания. То, что они также могут быть суперпоглотителями фруктозы, может означать, что они доставляют меньше фруктозы своим приятелям-кишечным бактериям. Подтвердить это предположение помогает исследование использования определенных штаммов «хороших бактерий» для лечения симптомов СРК.
Вернуться назад
Фруктоза в перспективе | Питание и обмен веществ
Фруктоза (сахароза или кукурузный сироп с высоким содержанием фруктозы (HFCS)) стала навязчивой идеей в питании. Окрашенные как однозначно вредные [1], даже как токсины, как в популярных [2], так и в научных публикациях [3-5], звучат призывы к регулированию и налогообложению со многих сторон. Хотя существует мало разногласий по поводу преимуществ снижения потребления сахара как средства ограничения калорийности или снижения общего количества углеводов, особенно для детей, лишь немногие исследования непосредственно проверяют эффект снижения фруктозы.Уникальные эффекты добавления фруктозы (по сравнению с глюкозой) менее очевидны (например, [6]) и должны быть согласованы с основными метаболическими процессами. По нашему мнению, обсуждение фруктозы демонстрирует недостаток сдержанности, напоминающий то, что сейчас признано поспешным суждением о пищевых жирах и холестерине. Кажется, стоит сделать шаг назад и взглянуть на фруктозу в перспективе.
С эволюционной точки зрения фруктоза, как и все углеводы, была знаком хороших времен. Обычно считается, что фоновая доступность углеводов и, в частности, фруктозы была низкой и непостоянной во времена палеолита.Колебания дефицита и изобилия рациона, вероятно, были намного больше, чем сейчас. Селективное преимущество заключалось в том, чтобы справиться с неурожаемыми годами, усвоить любую доступную низкую фоновую фруктозу, а также иметь возможность справиться с высокими уровнями потребления, которые часто сопровождаются внезапной удачей. Вполне вероятно, что то, что сегодня можно было бы считать чрезмерным потреблением, было частью нашего поведенческого репертуара на раннем этапе, и преимущество заключалось в накоплении запасов пищи, таких как жир и гликоген, которые могут быть недоступны в ближайшем будущем.Важно помнить, что эволюция не имеет ценности, и возможность съесть много за один присест может быть полезна в некоторых обстоятельствах. В общем, то, что называется патологией, — это человеческая точка зрения, а не истинная биологическая концепция; биологические системы выживают, приспосабливаясь к различным условиям.
В этом обзоре мы пытаемся описать, как метаболизм человека справляется с высоким содержанием фруктозы. Прием пищи сверх насущных потребностей в любом макроэлементе предполагает накопление в виде жира или гликогена, и высокое содержание фруктозы связано с обоими.Синтез гликогена, липогенез de novo (DNL) и синтез ТАГ, по сути, характерны для большого количества фруктозы. Фруктозу следует рассматривать в контексте прерывистого и сильно подкрепляющего источника пищи, а не в контексте яда.
Метаболизм фруктозы тесно связан с метаболизмом глюкозы (Обзоры: [7–10]). В этом сообщении мы пытаемся ответить на следующие вопросы: как можно совместить уникальные эффекты фруктозы, повышенного ТАГ и инсулинорезистентности, когда они наблюдаются, с непрерывностью метаболических путей глюкозы и фруктозы? Чем объясняются специфические эффекты парентерального введения фруктозы? Поскольку фруктоза и глюкоза связаны сходными путями, каков надлежащий контроль для эксперимента, в котором один сахар заменяется другим?
Наш вывод состоит в том, что фруктозу лучше всего рассматривать как часть общих путей углеводного обмена.Любые уникальные эффекты фруктозы опосредуются взаимодействием с глюкозой, а также значительным превращением фруктозы в глюкозу. Важное различие в процессинге двух сахаров заключается в ранних стадиях, гораздо более высоком сродстве фруктокиназы по сравнению с глюкокиназой, то есть регуляция находится на уровне контроля субстрата, влияя на популяцию триозофосфатов. Ожидается, что из-за их тесной связи замена диетической фруктозы глюкозой в качестве терапевтической меры будет варьироваться в зависимости от конкретных условий и индивидуальных реакций.Даже те исследования, которые показывают уникальные эффекты, как правило, имеют большие ошибки, и многие не могут различить эффекты добавленной фруктозы и глюкозы . Кроме того, некоторые экспериментальные исследования, возможно, большинство, требуют довольно высокого уровня фруктозы. Также ожидается, что замена фруктозы глюкозой будет менее эффективной в снижении факторов риска, чем замена любого углевода жиром. Ограничение углеводов в пище остается наиболее эффективной, хотя и недостаточно используемой стратегией против ожирения, диабета и метаболического синдрома в исследованиях различной продолжительности и протоколов ([11–18] и Дополнительный файл 1).
С точки зрения общественного здравоохранения, небольшое количество исследований, показывающих значительное улучшение метаболических нарушений за счет конкретного снижения содержания сахарозы или фруктозы в рационе, вызывает озабоченность, поскольку это рекомендуется для населения в целом. Было бы разумно избежать еще одного грандиозного эксперимента над всем населением — как была описана парадигма диеты и сердца — в отсутствие данных даже из небольших исследований и без учета непредвиденных последствий.
Общий взгляд на метаболизм фруктозы
«Да, в каталог вы попадаете для мужчин;
Как гончие и борзые, дворняги, спаниели, собачки,
Клещи, водяные коврики и полуволки убиты
Все по имени собак: ценный файл
Отличает быстрых, медлительных, тонких ,
Домработница, охотник, каждый
В соответствии с даром, который щедрая природа
Сверкнула в нем »
— Уильям Шекспир, Макбет
Фруктоза — это углевод.Он перерабатывается путем включения в метаболизм углеводов. Специфические эффекты, вызываемые повышением уровня фруктозы, происходят из-за увеличения промежуточных продуктов углеводного обмена. Для людей существует такой фон потребления пищи, при котором глюкоза почти всегда превышает фруктозу и где фруктоза быстро выводится, в то время как глюкоза поддерживается на постоянном уровне. Наиболее важно то, что значительное количество съеденной фруктозы превращается в глюкозу (рис. 1).
Рисунок 1Обзор основных аспектов метаболизма глюкозы и фруктозы в печени. Ключевые моменты, подчеркнутые в тексте: два сахара сходятся на уровне триозофосфатов (дигидроксиацетонфосфат (DHAP) и глицеральдегид-3-P (Ga-3-P)). Превращение фруктозы в триозофосфаты является однонаправленным, но фруктозо-1-фосфат является положительным эффектором глюкокиназы и регулирует синтез гликогена путем активации синтазы и ингибирования фосфорилазы. Последний эффект может быть разным у разных видов. Голубые стрелки показывают путь глюконеогенеза от фруктозы к глюкозе-6-P, которая, в свою очередь, может производить глюкозу или встраиваться в гликоген.
Метаболизм фруктозы и глюкозы сходится на уровне триозофосфатов (рис. 1). Основные проблемы метаболизма фруктозы — синтез глицерин-3-фосфата (глицерин-3-P) для синтеза триглицеридов, образование ацетил-КоА для цикла TCA и липогенез de novo (DNL) — происходят из этих промежуточных продуктов. Загадка для понимания метаболизма фруктозы заключается в том, как углерод в триозофосфатах знает, из фруктозы он или из глюкозы. Триозофосфаты также являются промежуточными продуктами глюконеогенеза; в нормальных условиях от 30 до 50% съеденной фруктозы превращается в глюкозу [8], а высокое содержание фруктозы также стимулирует выработку гликогена [19, 20].Наконец, глюкоза является основным стимулятором секреции инсулина, который контролирует метаболизм гликогена, сборку и распад триглицеридов, а также DNL. Взятые вместе, эти соображения предполагают некоторую осмотрительность при рассмотрении того, что может означать «добавленная фруктоза».
В такой сложной системе важными становятся экспериментальные детали. В частности, эффект от добавления фруктозы по сравнению с глюкозой в диету, фоновый состав которой уже содержит много углеводов, заметно отличается от того, в котором общее количество углеводов низкое.Специфические эффекты фруктозы, вероятно, представляют собой кинетические эффекты — более быстрое накопление промежуточных продуктов из фруктозы — но они также могут зависеть от индивидуальных условий.
При гиперкалорийной диете, когда уровень глюкозы в крови превышает высокий K m глюкокиназы, эффекты добавленной глюкозы или фруктозы будут неразличимы. McDevitt, et al. [21] , , например, обнаружили одинаковый уровень DNL у тучных и худых женщин, подвергшихся 50% перееданию глюкозы или сахарозы.При приеме пищи с низким содержанием сахара / углеводов выработка может контролироваться соединениями, получаемыми из фруктозы, но на самом деле было высказано предположение, что глюконеогенез может быть основным эффектом [7]. Сравнение низкого содержания фруктозы и глюкозы изучено недостаточно, по-видимому, потому, что они не считаются опасными для здоровья.
Какой контроль подходит для сравнения фруктозы и глюкозы?
Метаболизм не имеет ценности, то есть реакция на чрезмерное потребление фруктозы могла быть благоприятной для организма с прерывистым поступлением углеводов.Однако текущая дискуссия о питании состоит из хороших и плохих. Человек с метаболическим синдромом может уже плохо реагировать на широко изученные (и рекомендуемые) 55% углеводов, которые считаются стандартными [22, 23], но этот уровень немного выше, чем текущее потребление. В этих условиях любая добавленная фруктоза может быть хуже, чем глюкоза, но ожидается, что это будет незначительным нарушением эффекта высокого содержания углеводов, которое уже вызвало, например, высокий уровень триглицеридов.
В какой-то степени вопрос логичный. Многочисленные исследования продемонстрировали преимущества ограничения углеводов в целом (добавки). Тогда возникает вопрос, каков подходящий контроль для тех экспериментов, в которых сравниваются разные углеводы. Если есть нулевая гипотеза, то это то, что эффекты фруктозы в первую очередь связаны с ее ролью как углевода. Таким образом, подходящим сравнением были бы эксперименты, в которых все углеводы заменялись другим макроэлементом, обычно жиром.Мы приведем пример результатов исследований в лаборатории Джеффа Волека, которые можно сравнить с исследованиями фруктозы [12, 14, 24].
Поглощение глюкозы и фруктозы печенью
Печень — не единственный орган, который метаболизирует фруктозу, но печеночный метаболизм составляет не менее половины от общего количества и вызывает наибольшее беспокойство. Как своего рода командный центр в метаболизме, печень распределяет энергию по другим клеткам в виде глюкозы, лактата и триглицеридов, хотя другие ткани, кишечник, почки и мышцы, также могут напрямую перерабатывать фруктозу.В случае почек лактат и глюкоза из фруктозы также могут быть экспортированы.
Глюкокиназа, печеночная гексокиназа, имеет высокий K m — в диапазоне нормального уровня глюкозы в крови — так что печень и глюкоза крови находятся в равновесии. Уровень глюкозы в печени будет колебаться в зависимости от высокого уровня глюкозы в крови и других острых изменений, например, после голодания (рис. 1). В отличие от гексокиназ периферических тканей, глюкокиназа не подвержена ингибированию продукта глюкозой-6-P, но подлежит другой регуляции, включая повышение активности в ответ даже на небольшие количества фруктозы.
Фруктоза является плохим субстратом для глюкокиназы, и большая часть ее метаболизируется в результате реакции, катализируемой фруктокиназой, продуктом которой является фруктоза-1-P). Низкий K m фруктокиназы (0,5 мМ) означает, что фруктоза из плазмы быстро выводится. Фруктокиназа существует в двух изоформах, A и C. Последняя, экспрессируемая в основном в печени, имеет высокое сродство к фруктозе и приводит к быстрому включению и истощению АТФ, по крайней мере, как измерено у мышей. Фруктокиназа А имеет более широкое распределение в тканях, и ее более низкое сродство к фруктозе, по-видимому, снижает количество фруктозы для метаболизма в печени [25].Очевидная гетеротропная положительная кооперативность глюкокиназы, оказывается, связана с активацией фруктозой-1-P (рис. 1).
Регуляция печеночного процессинга фруктозы
Фосфорилирование фруктозы регулируется на уровне субстрата (низкий K m ) и транскрипции (экспрессия регулируется ChREBP (белок, связывающий элемент углеводного ответа) [26]). В самой реакции нет аллостерического или гормонального контроля, и обычно говорят, что фруктоза обходит фосфофруктокиназу-1 (PFK-1), которая регулирует метаболизм глюкозы.Кроме того, стадия лизиса, катализируемая альдолазой-B, отличается от альдолазы гликолизом, производящим дигидроксиацетонфосфат (DHAP) и глицеральдегид. Последние, в отличие от продуктов альдолазы, должны фосфорилироваться. Киназной реакции обычно уделяется мало внимания, но она указывает на роль печени как прямого потребителя фруктозы. Реакция альдолазы B обратима, а глицеральдегидкиназа — нет: «фруктонеогенеза» нет.
Хотя реакция фруктокиназы не подлежит аллостерической регуляции, сама стадия PFK-1 контролирует последующий метаболизм фруктозы.В условиях высокого энергетического заряда и высокого потребления фруктозы (в центре внимания в настоящее время), PFK-1 снижается с помощью АТФ и длинной петли обратной связи с цитратом. Точно так же фруктозо-1,6-бисфосфатаза стимулируется зарядом высокой энергии. Общий эффект — усиление глюконеогенеза за счет триозофосфатов (голубой путь на рисунке 1), чистое превращение фруктозы в глюкозу и, как описано ниже, высокий потенциал синтеза гликогена.
Регулирование поглощения и фосфорилирования
Фруктоза-1-P оказывает положительный аллостерический контроль глюкокиназы.Теперь понятно, что высокий K m глюкокиназы и ее очевидные аллостерические свойства обусловлены связыванием ингибирующего регулирующего глюкокиназу белка (RP; рис. 2), который снижает его сродство к субстрату. Фруктозо-1-фосфат снимает ингибирование путем связывания с RP, тем самым стабилизируя диссоциированные белки. Ингибирование является следствием транслокации комплекса глюкокиназа-RP в ядро [27]. Диссоциация RP и транспорт из ядра усиливается постпрандиальной глюкозой и инсулином [28].Дополнительная регуляция обеспечивается фруктозой-6-P в результате гликолиза, который действует как ингибитор обратной связи глюкокиназы, изменяя сродство к RP, восстанавливая связывание и возвращая фермент в ядро, тем самым создавая очевидное более высокое состояние K m .
Рисунок 2Контроль глюкокиназы. Связывание белка, регулирующего глюкокиназу (RP), снижает активность глюкокиназы (GK). Фруктоза-1-П снимает ингибирование, стабилизируя диссоциированный RP, в то время как F-6-P способствует ингибированию, стабилизируя комплекс.На рисунке не показано: комплекс GK-RP транспортируется в ядро [27]. Диссоциация приводит к транспорту из ядра.
Фруктозу можно рассматривать как метаболический сигнал для изобилия, требующий дополнительной глюкозы через облегчения ингибирования глюкокиназы. Этот ответ, очевидное уравновешивание добавленных фруктозы и глюкозы, может иметь общее значение. Введение только фруктозы может быть явно ненормальным состоянием для печени человека, которая развивалась в среде, где сахара всегда присутствовали вместе.Таким образом, фруктоза прямо или косвенно увеличивает эффективный уровень гликолитических промежуточных продуктов. Печеночный углеводный обмен реагирует на более низкие уровни фруктозы в плазме, чем на глюкозу, но ожидается, что комбинация фруктозы и глюкозы будет сильнее, чем одна фруктоза. Дальнейшее предсказание состоит в том, что в условиях высокого уровня глюкозы в плазме, в условиях, когда превышается K m глюкокиназы, различия между двумя сахарами должны исчезнуть. Опять же, DNL в условиях перекармливания соответствует этому прогнозу [21].
Глюконеогенез
Большая часть съеденной фруктозы превращается в глюкозу [8]. Деларю, и др. . [29] использовали дейтерированную глюкозу и природную 13 C-меченную фруктозу. Через 6 часов добавление 0,5 г / кг фруктозы привело к появлению 0,27 г / кг меченой глюкозы, тогда как при приеме внутрь 1 г / кг фруктозы было синтезировано 0,59 г / кг глюкозы. Эти значения представляют 56% и 59% нагрузки фруктозы. Аналогичные результаты, рассмотренные Саном [8], были получены другими.Обычно предполагается, что глюконеогенез фруктозы протекает, как показано голубыми стрелками на Рисунке 1 [8]. Не исключено, что глюконеогенез идет через пируват, хотя цикл ПЭП → пируват → ПЭП более энергетически невыгоден, чем обратная альдолазная реакция. Процесс также будет чувствителен к изменению энергетического заряда и присутствия других макроэлементов.
Гликоген
Как и ожидалось для реакции на время изобилия, высокое содержание фруктозы увеличивает запасы гликогена (рис. 1) либо непосредственно из глюкозы, либо из через глюконеогенез .Взаимодействие глюкозы и фруктозы изучали на первичных культурах гепатоцитов крыс, лишенных пищи в течение 24 часов. Парняк и Калант [19] измерили включение в гликоген [ 14 C] -глюкозы в присутствии холодной фруктозы и включение [ 14 C] -фруктозы в присутствии холодной глюкозы. Результаты, показанные на Фигуре 3, показывают, что метка любого из сахаров (в форме глюкозы) включается в гликоген дозозависимым образом.Каждый сахар усиливал включение другого (красные символы), и в обоих случаях высокий уровень инсулина (жирная линия) увеличивал общий выход по сравнению с его отсутствием (серая линия). Фруктоза приводит к большему включению, по-видимому, опять же из-за относительного более высокого поглощения и фосфорилирования, но наиболее заметным является сходство структур и степени превращения фруктозы в глюкозу.
Рисунок 3Включение меченой глюкозы или фруктозы в зависимости от концентрации меченого сахара и эффекта присутствия другого (холодного) сахара. Включение маркированного сахара при отсутствии другого показано синим символом и сплошной серой линией. Когда добавляется другой (немаркированный) сахар, происходит сдвиг во включении, как показано красным символом и серой пунктирной линией). Все эффекты усиливаются при добавлении инсулина (жирные черные линии). Рисунок перерисован из Parniak & Kalant [19].
Этот стимулирующий эффект фруктозы был продемонстрирован на людях Petersen, et al. [20] в элегантных экспериментах по измерению скорости синтеза гликогена в печени in vivo с использованием 13 C-ЯМР в условиях эугликемической гиперинсулинемии.Инфузии 13 C-меченой глюкозы в присутствии или в отсутствие фруктозы сопровождались холодной погоней для определения скорости синтеза и распада. В отличие от результатов с гепатоцитами, увеличение потока было связано с реакцией гликогенсинтазы (в 2,5 раза), а не с ингибированием активности фосфорилазы (рис. 1).
Инсулинорезистентность и метаболический синдром
Высокий уровень фруктозы часто описывается как причина инсулинорезистентности и метаболического синдрома (MetS), и есть несколько наблюдений, подтверждающих эту идею (например,г., [23]). Однако большинство таких экспериментов проводится в условиях высокого общего содержания углеводов, где, опять же, не уделяется должного внимания отдельным эффектам сахаров и их взаимодействиям. Кроме того, не все тесты показывают явный эффект фруктозы. Совсем недавно Lecoultre et al. [30] продемонстрировали снижение чувствительности печени к инсулину при переедании фруктозы. Однако, как видно на рисунке 4, наблюдались большие различия, и перекорм глюкозы имел аналогичный эффект.
Рисунок 4Чувствительность к инсулину печени (100 / (продукция глюкозы × инсулин натощак)). Субъекты мужского пола придерживались диеты для поддержания веса в течение 6–7 дней, а затем к той же диете было добавлено указанное количество сахара. Данные из ссылки [30].
Важным вопросом является рабочее определение инсулинорезистентности. Дирлевангер, и др. [31] измерял инсулин, необходимый для поддержания концентрации глюкозы 8 мМ во время гипергликемического зажима с добавлением или без добавления 16,7 мкМ / кг / мин фруктозы. Добавление фруктозы вызвало увеличение потребности в инсулине в 2 раза.Трехкратный и двукратный синтез гликогена. Используя радиоактивно меченую глюкозу, они показали, что фруктоза не влияла на выработку эндогенной глюкозы или общий выход по сравнению с контролем. Когда измерения проводились в соответствии с протоколом клэмп-теста с гиперинсулинемией и гипергликемией, добавление фруктозы вызывало увеличение общего выхода глюкозы на 11,1 мкМ / кг / мин, увеличивало чистое эндогенное производство глюкозы и увеличивало цикл глюкозы. Результаты были описаны как инсулинорезистентность печени, то есть для поддержания уровня глюкозы в крови в присутствии фруктозы требовалось больше инсулина, и, наоборот, высокий уровень инсулина не подавлял выработку глюкозы.
Хотя результаты соответствовали рабочему определению инсулинорезистентности, Dirlewanger, et al., l. [31] представили объяснение результатов, которое кажется нормальным ответом на экспериментальные условия и которое согласуется с механизмом на Рисунке 1: эффективная концентрация глюкозы в печени намного выше в группе фруктозы из-за обширного глюконеогенез. Это приведет к увеличению общего выхода глюкозы, несмотря на высокую концентрацию инсулина, концентрацию инсулина, которая была способна подавлять выход в контрольных образцах, содержащих только глюкозу.В то же время фруктоза-1-P, вызывая повышенную активность глюкокиназы, приведет к усиленному повторному захвату глюкозы и увеличению цикла глюкозы. Таким образом, очевидная инсулинорезистентность является результатом более высоких уровней промежуточных продуктов глюкозы в клетках, стимулированных фруктозой, и не представляет собой каких-либо основных пагубных изменений в физиологии печени.
Таким образом, концепция чувствительности к инсулину или резистентности к инсулину сильно зависит от метода измерения и рабочего определения. Резистентность к инсулину может быть характеристикой диабета 2 типа и sine qua non метаболического синдрома, но, в зависимости от того, что измеряется, не обязательно связана с дезадаптивными реакциями на пищу или другую стимуляцию, и я представляю продуктивную адаптацию к различным условиям.
Опять же, глядя на влияние общего количества пищевых углеводов в качестве контроля, известно, что ограничение углеводов по всем направлениям улучшит все характеристики МетС. Фактически было высказано предположение, что реакция всех индивидуальных маркеров MetS на снижение количества углеводов может служить рабочим определением [13], учитывая, что есть некоторый вопрос относительно того, существует ли синдром вообще [32]. Это важно в том смысле, что разумно, что, по крайней мере, некоторая часть инсулинорезистентности является понижающей регуляцией ответа из-за хронического высокого уровня инсулина, как и во многих гормональных системах.Снижение уровня глюкозы, а не фруктозы, снизит этот высокий уровень инсулина.
Критический проспективный тест был проведен Волеком и его коллегами, которые назначили 40 мужчин и женщин с избыточным весом и атерогенной дислипидемией, характерной для MetS (высокий уровень ТАГ, низкий уровень ХС-ЛПВП, высокая концентрация малых плотных ЛПНП), на кетогенную диету с очень низким содержанием углеводов. (VLCKD) в [12, 14, 24]. В качестве контроля использовалась диета с ограничением изокалорийности жиров. По сравнению с этими контрольными группами, группа VLCKD показала большее улучшение массы тела, гликемического контроля и многих характеристик MetS: TAG, HDL-C, апо B, апо A-1 и распределение частиц по размеру, а также распределение частиц LDL. больший противовоспалительный эффект.Степень, в которой эти эффекты ограничения углеводов были вызваны именно удалением фруктозы или сахарозы, неизвестна, но величина эффекта (хотя и в направлении улучшения), как правило, больше, чем наблюдаемые при сравнении глюкозы и фруктозы.
Дело в том, что мы знаем, что ограничение углеводов эффективно для уменьшения признаков метаболического синдрома. До тех пор, пока не появятся доказательства того, что был удален именно сахар, логика подсказывает, что ограничение углеводов является предпочтительным подходом.
Триглицериды
Гипертриглицеридемия, индуцированная углеводами, и ее обратная сторона, снижение уровня триглицеридов путем ограничения углеводов в пище, являются хорошо известными явлениями. Эффект обычно связывают с увеличением потока инсулина, который ингибирует липолиз, и снижением уровня глицерина-3-P, обеспечивающего субстрат для синтеза. Неизвестно, обладает ли фруктоза уникальным эффектом и в какой степени, но в нескольких статьях указывалось на высокий потенциал фруктозы по увеличению ТАГ в плазме по сравнению с эффектом глюкозы [22, 33–35].Однако имеются противоречивые сообщения [11], и, опять же, большинство недавних исследований было проведено относительно исходного уровня с высоким содержанием общих углеводов или необычно высоким содержанием фруктозы, или с обоими, что означает, что исходный уровень может быть уже очень высоким.
Чонг, и др. показал, что в острых постпрандиальных условиях малая активность радиоактивно меченной фруктозы проявляется в жирнокислотной части ТАГ, то есть жирные кислоты в основном происходят из эндогенного жира [35]. DNL относительно невелик для глюкозы или фруктозы.С другой стороны, основная часть (38%) глицерина происходит из меченой фруктозы, что согласуется с идеей о том, что основной эффект фруктозы заключается в более быстром или в большей степени заселении триозофосфатов.
На рисунке 5 показаны типичные результаты Stanhope, et al. [34], которые измерили относительные эффекты потребления напитков, подслащенных глюкозой или фруктозой, у субъектов с избыточным весом и ожирением. Напитки с сахаром обеспечивали 25% потребности в энергии в течение 10 недель.На рисунке показаны наложенные друг на друга кривые фруктозы и глюкозы из ссылки [34]. В среднем кривая фруктозы имеет большие отклонения после приема пищи, но разница в кривых составляет порядка 10%, что в сочетании с очень большими полосами погрешностей делает данные сомнительными. Планки погрешностей на рисунке фактически представляют стандартную ошибку среднего (SEM). SEM может предоставить статистическую меру различия в популяциях, но с точки зрения представления данных он не очень хорошо передает ощущение истинной изменчивости индивидуальных данных.Стандартное отклонение (SD), лучший индикатор истинного отклонения, получается путем умножения на √n, которое в данном случае составляет 3,7 для глюкозы и 4,1 для фруктозы. При таком большом разбросе пара выбросов изменит характер кривой. Другими словами, хотя фруктоза обладает уникальным действием, оно невелико и сильно варьируется.
Рисунок 5Триацилглицерин, циркулирующий в течение 24 часов до (черный) или через 2, 8 и 10 недель употребления подслащенных глюкозой (синий) или фруктозных напитков (красный), обеспечивая 25% энергии. Наложение рисунков 2A и 2B, перерисованных из Stanhope, et al. [34].
Аналогичным образом Teff, et al. [22] обнаружили значительно более высокий уровень ТАГ в группе, которая потребляла 30% своей общей энергии в напитках, подслащенных фруктозой, по сравнению с группой, употреблявшей напитки, подслащенные глюкозой. Этот уровень потребления довольно высок по сравнению со средним показателем в Соединенных Штатах (около 10%), но, опять же, большее значение имеют большие индивидуальные различия. Разделение субъектов в обеих группах на подгруппы на основе чувствительности к инсулину показало, что на самом деле различия из-за чувствительности к инсулину в пределах каждого сахара были столь же велики, как и различия между двумя сахарами.Как показано на фиг. 6, инсулинорезистентные субъекты в группе глюкозы имели более высокий ТАГ, чем чувствительные к инсулину субъекты в группе фруктозы. Из-за важности инсулинорезистентности разумно полагать, что инсулин будет реальной переменной, представляющей интерес.
Рисунок 6Сравнительные эффекты подслащенных фруктозой (красный) и глюкозосодержащих напитков (синий). Данные Teff, et al. [5] были объединены. Подгруппы, чувствительные к инсулину (- □ -) и резистентные к инсулину (- ○ -), были проанализированы отдельно от исходных популяций.
Связь между уровнями пищевых углеводов и ТАГ в плазме может быть единственным наиболее предсказуемым влиянием питательных веществ на метаболизм липидов (обзоры: [11, 13, 14]). То, что увеличение ТАГ в плазме не всегда наблюдается при кормлении с высоким содержанием фруктозы [36], предполагает, что это не главный или, по крайней мере, не единственный игрок. Надлежащий контроль, опять же, заключается в замене общих углеводов чем-то другим, особенно жирным. Хотя такого явного сравнения не проводилось, замена общих углеводов жирами всегда показывает гораздо большие изменения, чем эксперименты с обменом сахаров.Например, Волек, и др. . [24] сравнили две низкокалорийные диеты (~ 1500 ккал): диета с ограничением углеводов (% углевод: жир: белок = 12:59:28) и диета с низким содержанием жиров (LFD) (56:24:20) у 40 человек. субъекты с атерогенной дислипидемией (высокий уровень триглицеридов, низкий уровень ЛПВП, высокая концентрация малых плотных ЛПНП).
Рисунок 7 из Volek, et al. является репрезентативным. Изменения ТАГ от замены общих углеводов более значительны и надежны, чем исследования, в которых глюкоза заменяет фруктозу.Холленбек [36] провел обзор исследований, проведенных в 1993 г. по влиянию фруктозы на липидный обмен. По ее мнению, из 18 соответствующих исследований только 8 соответствовали критериям: 1) достаточный диетический и экспериментальный контроль, 2) глюкоза или крахмал для сравнения; и 3) имели ограниченную неоднородность в исследуемой популяции. Из этих 8 половина не показала изменений в ТАГ в плазме, в то время как одно не обнаружило изменений в нормальной группе, но увеличилось у субъектов с гипертриглицеридемией, и одно исследование аналогичным образом не обнаружило изменений в нормальной популяции с увеличением у субъектов с гиперинсулинемией.Только два исследования обнаружили увеличение ТАГ.
Фигура 7Влияние диеты на постпрандиальные липемические реакции у субъектов с атерогенной дислипидемией. Абсолютные значения ТАГ у субъектов, которые соблюдали кетогенную диету с очень низким содержанием углеводов (VLCKD) или диету с низким содержанием жиров (LF) в течение 12 недель. Перерисовано из Volek, et al. [24].
De novo липогенезDe novo липогенез (DNL) является важной характеристикой состояния с высоким энергетическим зарядом и / или высоким содержанием углеводов.Дифференциальные эффекты глюкозы и фруктозы (см. Ссылку [37]) следуют описанной здесь схеме. Например, Hudgins, et al. [38] измерил процентное увеличение пальмитата в VLDL-TAG. В соответствии с идеей о том, что прием фруктозы может изменить общую доступность сахаров в печени, они обнаружили, что фруктоза оказывает гораздо больший эффект, чем глюкоза, когда вводится отдельно; пероральный тест на толерантность к глюкозе (OGTT) имел небольшой эффект. Однако добавление глюкозы к дозе фруктозы увеличивает DNL, и когда общее количество углеводов становится высоким, превышая K m глюкокиназы, между сахарами мало различий (Рисунок 8), абсолютные изменения, однако, невелики и, опять же, существует большая изменчивость.
Рисунок 8Влияние сахара на процентное содержание пальмитата (16: 0) в ЛПОНП-ТГ до и после OGTT (глюкоза, 75 г; в среднем 0,8 г / кг) и указанные уровни фруктозы (F) и глюкозы ( ГРАММ). Взято из ссылки [38].
Одной из особенностей исследования Волека, описанного выше, было то, что, несмотря на 3-кратное увеличение потребления насыщенных жиров с пищей во время VLCKD, количество насыщенных жирных кислот в ТАГ и сложном эфире холестерина было значительно снижено по сравнению с контрольной группой с низким содержанием жира.То, что это было связано с уменьшением DNL, было показано соответствующим уменьшением пальмитолеиновой кислоты (16: 1n-7), продукта десатуразы. Пальмитолеиновая кислота присутствует в рационе только в низких концентрациях и обычно считается показателем DNL.
Эффекты чистой фруктозы
Из-за ограниченного воздействия на секрецию инсулина первоначально считалось, что фруктоза может быть желательным сахаром для людей с диабетом. Однако это оказалось не только неэффективным, но и привело к риску лактоацидоза.Подобные эффекты чистой фруктозы наблюдаются при парентеральном питании [39] или введении фруктозы во время упражнений [40]. Этот ответ традиционно объясняется кинетическим эффектом из-за быстрого фосфорилирования фруктозы и истощения АТФ. Одно из последствий — усиленный гликолиз и повышенное производство молочной кислоты. В условиях, когда доступны и фруктоза, и глюкоза, происходит несколько большее производство молочной кислоты из более высокого содержания фруктозы, хотя нет угрозы ацидоза, и молочная кислота является одним из способов, с помощью которых фруктоза снабжает энергией внепеченочные клетки.Хотя это предположение, разумный вывод состоит в том, что метаболизм в печени эволюционировал таким образом, что для метаболизма фруктозы требуется глюкоза. Цитозоль является гораздо более окислительным, чем митохондрия, и для превращения DHAP в глицерин-3-P требуется НАДН. При низком уровне НАДН глицеральдегид-3-П будет окисляться с образованием АТФ и НАДН, что приведет к превращению в пируват и выработку лактата. Кроме того, сообщалось о пищеварительных эффектах одной фруктозы, которые указывают на проблемы с абсорбцией.Возможно, результаты с одной фруктозой нельзя сравнивать с результатами, когда присутствуют оба сахара, и такие меры могут не быть хорошей моделью для потребления человеком, который почти никогда не включает чистую фруктозу.
Эффект энергетического заряда
Широко известно, что фруктоза истощает АТФ из-за фруктокиназной реакции [1], хотя это обычно наблюдается в изолированных культурах гепатоцитов и при добавлении чистой фруктозы. Основное внимание в настоящее время уделяется исследованиям, проводимым в условиях высокого энергетического заряда, а для реакций, характерных для фруктозы, — образования гликогена и триглицеридов — требуется АТФ.Veech, et al., , др. показал, что высокий уровень АТФ сопровождается диетой с высоким содержанием сахарозы [41]. Как ни странно, Абдельмалек, et al. [42] сообщил, что фруктоза снижает уровень АТФ в печени, что измерено с помощью 31 P-ядерного магнитного резонанса (ЯМР), но их данные не подтверждают этот вывод. Воздействие на АТФ может зависеть от конкретных условий. Влияние различных условий источника субстрата и, в частности, физических упражнений выходит за рамки этого обзора, но, опять же, существует значительная вариабельность, что видно даже по влиянию на скорость окисления различных сахаров (Обзор: [8]).Как отмечалось выше, присутствие фруктозы имеет тенденцию к увеличению уровней лактата в плазме, но, как правило, различия между диетической глюкозой и .