
Подъемы рук в стороны на тренажере
Займите исходное положение — Сидя в специальном тренажере, возьмите рукоятки, локти обоприте об валики.
- Сделайте вдох. Поднимите локти вверх.
- На выдохе опустите локти.
Подъемы рук в стороны на тренажере — выполнение
Данное упражнение прорабатывает средние дельты. Часто его рекомендуют выполнять начинающим атлетам по двум причинам:
- Оно не требует усилий для сохранения постоянного положения тела.
- Вероятность получения травмы очень низка.
Подъемы рук в сторону в тренажере — это упражнение, которое предназначено для изолированной проработки мышц плеч, а именно, среднего пучка. Так же в нем частичную нагрузку получает передний пучек. Если смотреть спереди или сзади, именно средний пучек дельт визуально делает плечи шире при виде.
Перед тем, как приступить к первому подходу, важно правильно настроить под себя тренажер. В отличии от упражнений со свободным весом, таких как махи с гантелями в стороны, где геометрия движения обусловлена положением ваших рук и туловища, при подъемах рук в стороны в тренажере, геометрия движения зависит от тренажера. Нужно выставить высоту сиденья так, чтобы движения ваших рук не были ограничены тренажером и вы могли работать на тренажере в полную амплитуду. Валики должны плотно прилегать к локтям с внешней стороны. Существует два вида тренажеров: без спинки, когда вы выполняете подъемы рук лицом к тренажеру, и со спинкой, когда вы выполняете движение спиной к тренажеру.
Некоторые спортсмены отмечают, что этот тренировки на этом тренажере являются для них неудобными. Такое возможно для людей ростом, отличным от среднего. Получается, что движение рук не совпадают с траекторией движения упоров тренажера. В таком случае, подъемы рук в стороны в тренажере можно заменить похожи упражнением со свободным весом, таким как махи гантелями в стороны.
Раздел:
Другие упражнения на плечи:
Как выполянть: 3-4 подхода по 10-20 повторений.
МАХИ В ТРЕНАЖЕРЕ — УПРАЖНЕНИЕ ДЛЯ ТРЕНИРОВКИ СРЕДНЕЙ и ЗАДНЕЙ ДЕЛЬТ, ПРАВИЛЬНАЯ ТЕХНИКА ВЫПОЛНЕНИЯ МАХОВ В ТРЕНАЖЕРЕ
Махи в тренажере – это формирующее упражнение для тренинга средней дельты плечевого пояса, эмитирующее махи с гантелями стоя. Махи в тренажере можно использовать начинающим атлетам, которые ещё не умеют правильно выполнять более сложные технически упражнения, поскольку, если новичку некому поставить технику, то упражнения со свободными весами они иногда выполняют просто с умопомрачительной фантазией. Другими словами, плюсом упражнения является то, что в нем атлет вынужден выполнять движение в заданной амплитуде. Конечно, опытные качата могут быть, наоборот, недовольны этим фактом, поскольку умеют подбирать под себя более адекватные углы нагрузки, но и им может пригодиться тренажер для плеч.
Махи в тренажере можно использовать начинающим атлетам, которые ещё не умеют правильно выполнять более сложные технически упражнения, поскольку, если новичку некому поставить технику, то упражнения со свободными весами они иногда выполняют просто с умопомрачительной фантазией. Другими словами, плюсом упражнения является то, что в нем атлет вынужден выполнять движение в заданной амплитуде. Конечно, опытные качата могут быть, наоборот, недовольны этим фактом, поскольку умеют подбирать под себя более адекватные углы нагрузки, но и им может пригодиться тренажер для плеч.
Махи в тренажере позволяют качественно сосредоточиться на работе целевой мышечной группе, поскольку атлету не приходится отвлекаться на удержание штанги или гантелей. Да, заданная амплитуда движения, конечно, может быть не подходящей для атлетов с каким-то особенным строением скелета, но большинству тренажер подойдет. Именно поэтому, если Вы хотите прокачать дельты, скажем, наполнить их хорошо кровью в конце тренировки, то махи в тренажере – это не худший выбор. Единственное, что Вам следует помнить, это то, что рабочей дельтой всегда является та, которая находится над осью плечевого сустава. Именно поэтому опускать локти ни в коем случае нельзя, поскольку передний пучок дельт руку в сторону отводить не может, а средний, находясь под осью плечевого сустава, иннервировать невозможно, в таких условиях нагрузку на себя берут широчайшие мышцы спины.
Единственное, что Вам следует помнить, это то, что рабочей дельтой всегда является та, которая находится над осью плечевого сустава. Именно поэтому опускать локти ни в коем случае нельзя, поскольку передний пучок дельт руку в сторону отводить не может, а средний, находясь под осью плечевого сустава, иннервировать невозможно, в таких условиях нагрузку на себя берут широчайшие мышцы спины.
Основными работающими мышечными группами во время выполнения махов в тренажере являются дельты, причем, не только средняя, но ещё и задняя, только для этого нужно немного наклониться вперед. Само собой, что иннервируется так же и трапециевидная мышца, а мышцы брюшного пресса и длинные мышцы спины выполняют функцию стабилизаторов. Вообще, не страшна иннервация никаких мышечных групп, кроме широчайших мышц спины. Широчайшие мышцы могут отбирать нагрузку у дельт и будут это делать, если Вы опустите локти ниже кисти, поскольку в таком положении иннервировать плечи сложнее, а широчайшие мышцы спины легче, при этом, широчайшие сами по себе сильнее дельт, так что они с радостью «украдут» всю нагрузку.
Работающим суставом является только плечевой, поэтому упражнение изолирующее, но это не плохо, просто это надо учитывать. Суть в том, что формирующие упражнения просто не надо выполнять в силовом стиле, их нужно выполнять с умеренным весом, с которым атлет может достичь позитивного отказа на 8-15 повторении. Если же Вы попробуете использовать более тяжелые веса, то в связи с тем, что в нижней фазе амплитуды движения в сидячем положении «читинг» невозможен, Вы просто сместите нагрузку в широчайшие мышцы спины, опустив локти, или не выполните упражнение вообще. Впрочем, если у Вас есть тренировочный партнер, тогда он может Вам помогать в нижней фазе, либо можно работать в укороченной амплитуде, но использовать такие методы угнетения мышечных волокон стоит уже на продвинутом уровне.
1) Сядьте в тренажер, немного наклонив корпус вперед, чтобы средняя и задняя дельты оказались над осью плечевого сустава.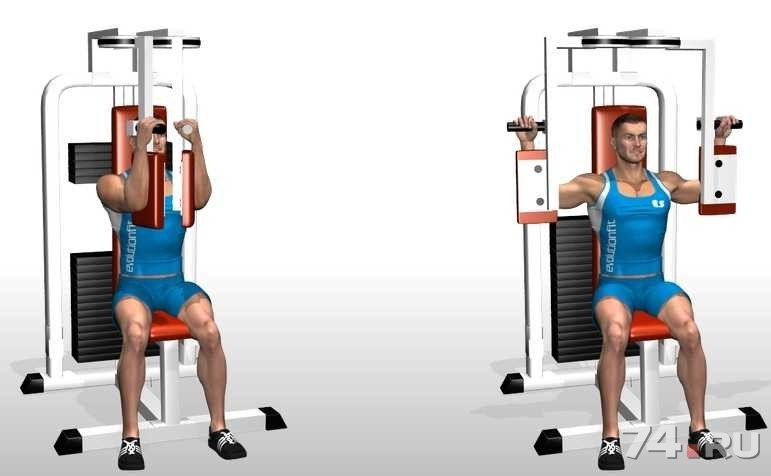
2) Возьмитесь за рукоятки тренажера и поднимите локти выше кистевых суставов, чтобы исключить из работы широчайшие мышцы спины.
3) Вдохните воздух и на выдохе мощным толчком разведите руки в стороны, чтобы в верхней точке амплитуды движения локти оказались перпендикулярно полу.
4) Немного задержавшись в точке пикового сокращения дельт, медленно и подконтрольно вернитесь в исходное положение.
Махи в тренажере – примечания
1) Во время выполнения махов ни в коем случае не выпрямляйте корпус, поскольку это сместит нагрузку с задней дельты и, в лучшем случае, Вы прокачаете только средний пучок дельтовидных мышц плечевого пояса.
2) Не поднимайте локти выше плечевого сустава, поскольку в этой фазе амплитуды движения дельты уже не способны брать на себя нагрузку.
3) Ни в коем случае не опускайте локти ниже кистевых суставов, поскольку это полностью нивелирует весь смысл упражнения.
4) Опускайте руки вниз медленно и подконтрольно, а позитивную фазу движения выполняйте мощно и быстро.
Плечевой пояс наделен очень подвижным суставом, который способен крутиться буквально во все стороны, поэтому дельты состоят аж из трех головок, выполняющих совершенно разные функции. Уникальна эта мышца ещё и тем, что каждая её головка отделена от другой мышечной фасцией, что, по сути, делает каждую из них самостоятельной мышцей. Передний пучок дельт отвечает за отведение рук от корпуса, поэтому он активно участвует во всем жимах, задняя дельта отвечает за притягивание рук к корпусу, а средняя является подсобной и иннервируется практически всегда, но выполняет лишь функцию стабилизатора, поэтому отдельно её так же нужно прокачивать. Собственно, то, какая дельта будет работать, зависит от того, какая из них находится над осью плечевого сустава – это ключевой момент!
Подводя итоги, можно сказать, что махи в тренажере являются неплохим формирующим упражнением для прокачки задней и средней дельт плечевого пояса, которое могут использовать атлеты разного уровня подготовки. Ключевым моментом является положение дельт относительно оси плечевого сустава, поскольку рабочей является та, которая находится над ней. Вторым, не менее важным, но более легко соблюдаемым, правилом является положение локтя относительно кисти, а именно то, что локоть всегда должен быть выше кисти. Поднимать локти выше плечевого пояса не нужно, поскольку в этом положении будут работать уже не дельты, а трапециевидная мышца, что является пустой тратой времени, поскольку прокачать её таким же маленьким весом, как плечи, просто не получится!
Ключевым моментом является положение дельт относительно оси плечевого сустава, поскольку рабочей является та, которая находится над ней. Вторым, не менее важным, но более легко соблюдаемым, правилом является положение локтя относительно кисти, а именно то, что локоть всегда должен быть выше кисти. Поднимать локти выше плечевого пояса не нужно, поскольку в этом положении будут работать уже не дельты, а трапециевидная мышца, что является пустой тратой времени, поскольку прокачать её таким же маленьким весом, как плечи, просто не получится!
Упражнения для бодибилдинга
Разведение рук в тренажере (обратные разведения) техника
Разведение рук в тренажере (обратные разведения) – изолирующее упражнение для дельтовидных мышц, похожее по механике на махи гантелями в наклоне. Особенностью упражнения можно считать возможность упора туловища в скамью тренажера, за счет чего происходит качественная проработка мышц – корпус лишается возможности к раскачиванию.
Работающие мышцы и способы хвата
Обратные разведения напрямую воздействуют на дельтовидные мышцы (а именно – на задние пучки). Дополнительно включаются:
- трапециевидная мышца;
- ромбовидная.
Обратные разведения в тренажере могут выполняться с использованием различных вариантов хвата:
- классический, наиболее распространенный вид хвата – когда ладони находятся на вертикальных ручках тренажера друг напротив друга, а большие пальцы направлены в потолок;
- другой вид хвата противоположен классическому – кисти рук направлены друг на друга тыльной стороной, большие пальцы обращены к полу, хват осуществляется за вертикальные ручки;
- последний вариант хвата является специфичным, так как на большей части тренажеров отсутствуют ручки для такого вида хватов – хват осуществляется за ручки, которые расположены горизонтально, большие пальцы направлены друг на друга, а ладони обращены вниз.
Исходное положение
Прежде чем приступать к выполнению упражнений на разведение рук, потребуется регулировка некоторых параметров тренажера. Настройте его так, чтобы, находясь в расслабленном состоянии, ваши руки на рукоятках располагались примерно на ширине плеч. При полном отведении рук они должны быть практически прямыми и максимально параллельными полу.
Настройте его так, чтобы, находясь в расслабленном состоянии, ваши руки на рукоятках располагались примерно на ширине плеч. При полном отведении рук они должны быть практически прямыми и максимально параллельными полу.
Непосредственно в ходе выполнения упражнения следите за расположением собственного тела: грудная клетка должна плотно прижиматься к спинке тренажера, корпус расположен строго перпендикулярно полу, спина имеет небольшой прогиб в области поясницы, руки прямые, ладони располагаются друг напротив друга. До начала выполнения упражнения на разведение рук в тренажере немного отодвиньте друг от друга рукоятки, чтобы слегка приподнять с упоров вес.
Техника выполнения
Перед моментом подъема груза выполните глубокий вдох и на некоторое время задержите дыхание. Затем, ощущая напряжение задних дельт, разведите рукоятки на максимально возможное расстояние назад. Следите, чтобы локти отодвинулись за плоскость спины. Если вы хотите максимально нагрузить именно дельтовидные мышцы и выключить трапецию, старайтесь разворачивать плечи вперед.
В то время, когда руки достигнут максимально возможного отведения, задержите их ненадолго в этом положении, продолжая удерживать напряжение мышц, после этого можете выполнить выдох и вернуться в исходное положение. Затем на секунду задержитесь в начальном положении и продолжайте выполнять повторы упражнения.
https://www.youtube.com/watch?v=02PapkNQQwU
Некоторые тренажеры сконструированы таким образом, что выполнять обратные разведения с полностью прямыми руками не представляется возможным, поэтому допускается выполнение со слегка согнутыми руками. Основным условием является неподвижность локтевого сустава – при осуществлении разведения он не должен двигаться.
Рекомендации
Для облегчения работы при выполнении упражнения на отведение рук нужно представить себе, что вы отводите назад не рукоятки тренажера, а собственные локти. Таким образом получится исключить из работы руки и больше задействовать именно целевые мышцы.
Корпус во время выполнения упражнения должен быть максимально неподвижен, старайтесь фиксировать его в одном положении.
Это обеспечит большую эффективность выполнения и безопасность для вас. При выполнении разведения рук произвести более мощное усилие позволит правильное дыхание.
Для максимального включения в работу дельтовидных мышц старайтесь в ходе выполнения упражнения по максимуму отводить локти за спину. Если сделать этого вы не можете, это означает, что нужно немного уменьшить вес снаряда или же развивать большую гибкость плечевых суставов.
Тренировка для мужских плеч, на которые можно взвалить абсолютно все
Красивые, мощные и широкие плечи — визитная карточка настоящего мужчины.И чтобы они такими стали, тренировка на плечи должна быть не только интенсивной, но и технически безупречной, так как травмы именно этих суставов подстерегают на каждом шагу. Поэтому подробно разбираем технику.
Жим сидя (армейский)
Цель: мышцы плечевого пояса, трицепс.
Техника выполнения:
● Сядьте удобно на скамью: упражнение можно выполнять как на скамье со спинкой, так и на скамье без спинки; со свободным весом или в тренажёре.
● Удержите вес в руках на уровне подбородка. Держите локти перед собой, не разводя их в стороны.
● Разгибая руки, вытолкните снаряд вверх.
Ошибки:
● Не делайте рывковое движение до жёсткого замыкания локтевого сустава при выпрямлении рук.
● Не сутультесь и не заводите плечи вовнутрь.
● Не разводите локти.
Жим Арнольда
Цель: мышцы плечевого пояса (передняя и средняя дельты).
Техника выполнения:
● Сядьте удобно на скамью.
● Удержите гантели на согнутых руках перед грудью.
● Разведите локти в стороны с последующим распрямлением рук и толчком гантелей вверх.
● По той же траектории вернитесь в исходное положение.
Ошибки:
● Не прижимайте локти к туловищу — удерживайте гантели навесу.
● При выпрямлении рук избегайте жёсткого замыкания локтевого сустава.
● Не отводите гантели назад. Толчок гантелей должен быть вверх или немного вперёд.
Махи через стороны сидя на скамье
Цель: мышцы плечевого пояса (средняя дельта, трапеции).
Техника выполнения:
● Сядьте удобно на скамью, выпрямив спину.
● Выполните подъём рук через стороны до образования прямой линии между руками и плечевым поясом (до параллели с полом).
Ошибки:
● Не разворачивайте кисти рук наружу, так вы перенесёте нагрузку со средней на переднюю дельтовидную мышцу.
● Не выполняйте упражнение с большим весом.
Швунги со штангой
Цель: мышцы плечевого пояса.
Техника выполнения:
● Поднимите штангу с пола и удержите её руками на верхнем отделе грудных мышц
● Выполните подсед и инерционным движением, разгибая ноги, вытолкните снаряд вверх.
● В финале движения следите за равновесием. избегайте опрокидывания назад.
Можете работать и с большими весами, так как при правильном выполнении упражнения нагрузка распределяется на всё тело.
Ошибки:
● Не выполняйте упражнение только за счёт силы рук. Движение вверх должно быть инерционным, импульс которому задают ноги.
● Избегайте жесткого замыкания локтевого сустава.
● Не запрокидывайтесь назад.
Высокая тяга к подбородку (с ЕZ грифом)
Цель: мышцы плечевого пояса (трапеции, передняя и средняя дельты).
Техника выполнения:
● Встаньте, установив ноги на комфортной ширине.
● Опустите снаряд перед собой на прямленных руках.
● Сгибайте руки к подбородку, подтягивая снаряд.
Ошибки:
● Не заваливайте локти вниз во время движения. В финале упражнения они должны быть направлены строго вверх по наклонной.
● Не выполняйте движение за счёт силы бицепсов, концентрируйтесь на плечах.
 youtube.com/embed/Jb1h5bQngVo?iv_load_policy=3&modestbranding=1&rel=0&autohide=1&playsinline=1&autoplay=0″ sandbox=»allow-scripts allow-same-origin allow-popups»/>
youtube.com/embed/Jb1h5bQngVo?iv_load_policy=3&modestbranding=1&rel=0&autohide=1&playsinline=1&autoplay=0″ sandbox=»allow-scripts allow-same-origin allow-popups»/>
Шраги
Цель: трапеции.
Техника выполнения:
● Встаньте, установив ноги на комфортной ширине.
● Опустите снаряд перед собой на прямленных руках.
● Выполните движение плечами вверх, при этом сохраняя руки прямыми. Они должны выполнять только роль держателей.
Ошибки:
● Не сгибайте руки, движение должно быть только плечами вверх, к ушам.
Махи через стороны в наклоне
Цель: мышцы плечевого пояса (задняя дельта, трапеции, середина спины).
Техника выполнения:
● Встаньте, установив ноги на комфортной ширине и наклонитесь.
● Удержите вес на опущенных вниз выпрямленных руках.
● Выполните отведение с подъёмом рук через стороны, до параллели с полом.
Ошибки:
● Не сгибайте локти и не заводите их за спину, так вы перенесёте нагрузку на широчайшие мышцы спины.
● Не делайте рывковых движений и не работайте с большим весом.
Махи одной рукой в кроссовере
Цель: мышцы плечевого пояса (передняя, средняя дельта).
Техника выполнения:
● Расположитесь рядом с кроссовером или другой блочной тягой.
● Выполните отведение прижатой руки от тела в сторону.
Ошибки:
● Не сгибайте сильно руку в локте.
● Не выполняйте движение рывком с запрокидыванием тела назад.
Примерная программа №1:Жим армейский
Махи через стороны
Высокая тяга
Махи в наклоне
Шраги
Примерная программа №2:Швунги
Жим Арнольда
Махи в наклоне
Махи в кроссовере через стороны
Шраги
Как работать с программой:1. Если тренируетесь с приоритетом на плечи — занимайтесь по программе 2 раза в неделю.
2. Если тренируетесь в сплите – занимайтесь по программе 1 раз в неделю.
3. Вес и количество повторений подбирайте в зависимости от целей.
(1142)
comments powered by HyperComments
|

Махи руками назад на нижнем блоке в наклоне
Описывается техника выполнения махов руками назад на нижнем блоке в наклоне, тренируемые мышцы, аналоги упражнения, нюансы выполнения махов и обучающее видео.
Тренируемые мышцы
Главную нагрузку получают задние дельты, дополнительную получат трапециевидная и ромбовидная мышцы.
Описание упражнения
В процессе всего движения спина остаётся на одном месте и ровная, когда руки разведены, локти смотрят в потолок, а не назад. В верхней точке старайтесь не сдвигать лопатки друг к другу и не поднимайте плечи, работают только плечи, иначе львиная доля нагрузки уйдёт в трапецию. На протяжении всего упражнения, локти слегка согнуты.
Аналоги упражнения
— Махи руками назад в кроссовере
— Разведение гантелей в стороны в наклоне
Нюансы выполнения махов
1) Поднимайте руки так, чтобы локти были не выше плеч, выше не нужно, дальше Вам придётся сводить лопатки и тогда нагрузка из задние дельты уйдёт в трапецию и ромбовидную мышцу.
2) Когда нагнули спину, обязательно слегка согните ноги в коленях, это снимет напряжения в пояснице и позволит держать её прогнутой и прямой.
3) Спина должны быть под одинаковым углом наклона, как в верхней, так и в нижней точки упражнения.
4) Махи можно выполнять одновременно двумя руками или одной, как стоя, так и сидя на гимнастической скамье. Одно отличие, что сидя проблематично прогнуть спину до параллели с полом, что в положении стоя это сделать более реально.
5) Локти всегда должны быть немного согнутыми, это снимет не только нагрузку на локти, но и сконцентрирует всю нагрузку на заднем пучке дельт и помните локти смотрят строго вверх.
Упражнения на заднюю дельту
1) Разведение гантелей в стороны в наклоне
2) Махи руками назад в кроссовере
3) Разведение рук в тренажёре назад
Махи руками назад на нижнем блоке в наклоне видео
Рекомендуем Вам:
Метки: упражненияРазведение гантелей в стороны. Подъем гантелей через стороны. Махи руками с гантелями.
Из-за своего строения, фактически не существует какого-то одного упражнения, которое бы могло нагрузить плечо равномерно и полностью. Тренировка плеч должна включать различные упражнения, направленные на разные пучки мышцы. Выполняя подъемы гантелей через стороны вы максимально сильно нагрузите средние пучки дельтовидных мышц, что сделает ваши плечи визуально более массивными и объемными.
Варианты выполнения упражнения
Ниже мы покажем все способы и технику выполнения подъемов гантелей через стороны.
Разведение гантелей в стороны стоя
Возьмите гантели нейтральным хватом ладонями друг к другу. Встаньте прямо, слегка согните ноги и немного наклонитесь корпусом вперед. Руки немного согнуты в локтях. На выдохе разведите гантели в стороны не меняя угол в локтях. На вдохе опустите руки в исходное положение.
Выполняйте подъем так, чтобы в верхней точке движения плечо, локоть и кисть образовывали одну прямую линию. Старайтесь как можно меньше раскачиваться туловищем — инерция значительно уменьшает эффективность упражнения. Не стоит использовать слишком большой вес, иначе вы просто не сможете почувствовать работу дельтовидных мышц. На вдохе — плавно вниз, на выдохе — вверх.
Разведение гантелей в стороны сидя
Сядьте на скамью и возьмите гантели нейтральным хватом ладонями друг к другу. Руки немного согнуты в локтях. На выдохе разведите гантели в стороны не меняя угол в локтях. На вдохе опустите руки в исходное положение.
Чтобы движения были четче, используйте не слишком большой вес — так тренировка средней части дельтовидной мышцы будет эффективнее. Это важно на начальном этапе занятий. Для большей результативности в верхней точке задерживайте руки на несколько секунд.
Отведение одной руки с гантелью в сторону
Возьмите в одну руку гантель, другой рукой крепко держитесь за неподвижную опору. На выдохе отведите рабочую руку в сторону. На вдохе вернитесь в исходное положение. Сделайте необходимое количество раз и повторите упражнение для другой руки.
Для большей эффективности не следует в нижней точке расслаблять дельтовидные мышцы и опускать гантели на бедра. Держите их на некотором расстоянии, чтобы нагрузка плечи была на протяжении всего упражнения.
Видео
Почему мы размахиваем руками во время ходьбы?
Вы когда-нибудь задумывались, почему мы качаем руками во время ходьбы?
Это любопытное явление, поскольку руки не играют очевидной роли в передвижении. Вы все еще можете нормально ходить, когда у вас руки полны продуктов или руки в карманах, так зачем мы это делаем? Разве это не пустая трата энергии?
Два независимых исследования, оба опубликованные в 2009 году, могут дать некоторые ответы на эту загадку.
Ответ…
Используя комбинацию механических моделей, компьютерного моделирования и тестов с участием людей, ходящих и бегунов, исследования пришли к выводу, что раскачивание руки просто … происходит. Другими словами, вам не нужны мышцы, чтобы ваши руки начали раскачиваться, хотя вы можете использовать мышцы, чтобы махать руками выше или сильнее.
Если вы идете, и ваши руки расслабленно свисают по бокам, они обычно будут качаться синхронно с противоположной ногой. То есть ваша правая рука поворачивается вперед, когда левая нога делает шаг вперед, и наоборот.
Раскачивание рук и шаг
Но хотя это происходит пассивно, раскачивание руками играет важную роль в повышении эффективности вашего шага. Это достигается за счет уравновешивания туловища и бедер и предотвращения их чрезмерного скручивания и покачивания.
Удерживание рук по бокам во время ходьбы требует на двенадцать процентов больше энергии, чем если бы ваши руки могли раскачиваться естественным образом. Принуждение рук к движению синхронно с ногой на той же стороне тела требует на двадцать шесть процентов больше энергии, чем при обычной ходьбе.
Даже когда у ходоков были привязаны руки к бокам и не требовалось никакого мышечного контроля, чтобы удерживать их в неподвижном состоянии, они использовали на семь процентов больше энергии, чем когда их руки могли свободно раскачиваться. Это говорит о том, что размахивание руками — это не просто пассивный побочный продукт ходьбы, но фактически облегчает ходьбу.
Подробнее: Динамическое раскачивание руки при ходьбе человека (PDF) и Управление и функция раскачивания руки при ходьбе и беге человека (PDF)
(PDF) Влияние качания руки на эффективную энергию во время вертикальных прыжков: экспериментальное и моделирование
18
Ссылки
Эшби Б.М., Delp SL.Моделирование оптимального управления раскрывает механизмы, с помощью которых движение руки
улучшает выполнение прыжков в длину с места. J Biomech 2006: 39: 1726-1734.
Бобберт М.Ф., Касиус Л.Дж. Связано ли влияние встречного движения на высоту прыжка с развитием активного состояния
? Med Sci Sports Exerc 2005: 37: 440-446.
Bobbert MF, Casius LJ, Sijpkens IW, Jaspers RT. Люди приспосабливают контроль к начальной глубине приседа
в вертикальных приседаниях. J Appl Physiol 2008: 105: 1428-1440.
Bobbert MF, van Soest AJ. Почему люди прыгают именно так? Exerc Sport Sci Rev 2001:
29: 95-102.
Cheng KB, Wang CH, Chen HC, Wu CD, Chiu HT. Механизмы, которые позволяют движению руки
для повышения производительности вертикального прыжка — исследование моделирования. J Biomech 2008: 41: 1847-1854.
Дапена Дж., Чанг CS. Вертикальные и радиальные движения тела на отрывном этапе
прыжки в высоту. Med Sci Sports Exerc 1988: 20: 290-302.
de Looze MP, Kingma I, Bussmann JBJ, Toussaint HM.Валидация модели сегментов
с динамической связью для расчета шарнирных моментов в подъемах. Clin Biomech 1992: 7: 161-169.
Domire ZJ, Challis JH. Анализ индуцированной энергии для определения механизма повышения производительности
в результате взмаха руки во время прыжка. Sports Biomech 2010: 9:
38-46.
Эбаши С., Эндо М. Ион кальция и сокращение мышц. Prog Biophys Mol Biol 1968: 18:
123-183.
Фельтнер М.Э., епископ Е.Дж., Перес К.М.Сегментарный и кинетический вклады в вертикальных прыжках
с махом руки и без него. Res Q Exerc Sport 2004: 75: 216-230.
Фельтнер М.Э., Фрашетти Д.Д., Крисп Р.Дж. Увеличение верхней конечности нижней конечности
кинетика при вертикальных прыжках с встречным движением. J Sports Sci 1999: 17: 449–466.
Хара М., Шибаяма А., Такешита Д., Фукасиро С. Эффект замаха руки на нижних
конечностях при вертикальных прыжках.J Biomech 2006: 39: 2503–2511.
Харман Э.А., Розенштейн М.Т., Фрикман П.Н., Розенштейн Р.М. Влияние рук и противодействия
на вертикальные прыжки. Med Sci Sports Exerc 1990: 22: 825-833.
Хатце Х. Модель миокибернетического контроля скелетных мышц. Биол Киберн 1977: 25: 103-119.
Халид В., Амин М., Бобер Т. Влияние движения верхних конечностей на отрыв в
вертикальном прыжке. В: Tsarouchas I, Terauds J, Gowitzke B, Holt L, ред.Биомеханика в спорте
V. Афины: Греческий научно-исследовательский институт спорта, 1989: 375-379.
Кистемейкер Д.А., Ван Суст А.Дж., Бобберт М.Ф. Возможен ли контроль точки равновесия для быстрых целенаправленных односуставных движений
? J Neurophysiol 2006: 95: 2898–2912.
Сочетание движений старта: взмах руки
В предыдущих статьях Race Club мы обсуждали важность взаимосвязанных движений во всех четырех гребках и то, насколько сильно они могут влиять на тягу и скорость пловца.Связанные движения старта так же важны, как и в каждом из четырех ударов.
На старте есть три основных движения сцепления; подъем головы, движение руки и подъем задней ноги в воздух. На этой и прошлой неделе в подписке на дорожках 2, 3 и 4 в рамках нашей стартовой серии мы выделяем движение руки олимпийца Брэда Тэнди, у которого один из лучших стартов в мире. Движение руки на старте может быть мощным сцепным движением.
Хотя есть некоторые вариации следующих техник, мы видим в основном четыре разных движения рук у пловцов, когда они покидают блок перед тем, как занять обтекаемую позицию для входа.Поскольку количество кинетической энергии в движении руки — это то, что влияет на движение ног и рук, я ранжирую четыре различных движения рук от самой высокой энергии до самой низкой. Другими словами, они ранжируются от того, что мы считаем лучшим движением руки к худшему.
- Большой круг . Выпрямляя руки из согнутого положения, пловец в четыре раза увеличивает кинетическую энергию этого кругового движения. Ключ состоит в том, чтобы держать руки прямыми на протяжении всего кругового движения и, выгнув спину пловца в воздухе, продолжить круг руками высоко над верхней частью спины пловца, образуя большой круг в обтекаемом положении.Брэд делает это движение лучше, чем кто-либо из тех, кого я когда-либо видел.
- Малый круг. За счет сгибания локтей и создания непрерывного круга меньшего размера кинетическая энергия движения руки уменьшается. Эта техника сгибания локтей, которую используют большинство элитных американских пловцов, проще и безопаснее, но может не дать такого хорошего старта, как большое круговое движение руки. Некоторые тренеры называют эту технику «быстрым стартом».
- Движение возрастных групп. Я вижу много молодых пловцов, использующих эту технику.В этом старте пловец начинает движение прямой рукой по круговой схеме, но останавливает движение, как только руки достигают ног пловца. Затем пловец медленно меняет направление руки под телом обратно в обтекаемое положение. Прекращая движение рук, пловец теряет ценную энергию сцепления.
- Передний ход рычага. При использовании этой техники пловец просто освобождает руки от блока и медленно переводит их вперед, принимая плавное положение.Это медленное движение рычагов дает очень небольшую кинетическую энергию и минимальный эффект связи.
Мы смогли обучить старт Тэнди, используя большую прямую руку, круговое движение руки, конкурентоспособных пловцов всех возрастов, включая мастеров. Фактически Брэд начал использовать эту стартовую технику в возрасте семи лет. Используя прогрессивную технику, мы начинаем с моделирования засушливых земель и продвигаемся к стартовым блокам. Большинству пловцов удается добиться правильного движения рук за несколько попыток.
При сравнении американского рекордсмена Зейна Гроте, использующего технологию измерителя скорости, мы обнаружили, что он был на 2% быстрее на старте с 12 метров, используя движение руки Тэнди по большому кругу по сравнению с движением руки меньшего круга. Это означало, что он был на 0,1 секунды быстрее, что было достаточно, чтобы выиграть или проиграть гонку.
Как и в любой другой технике плавания, пловцы должны практиковаться, практиковаться и тренироваться, чтобы совершенствовать ее. Мы рекомендуем попрактиковаться в этом старте 100 раз, прежде чем пробовать его в гонке.Как только вы его опустите, вы почувствуете, что летите по воздуху.
Мы надеемся, что вам понравятся наши недавние веб-эпизоды по нашей подписке, поскольку мы разбиваем некоторые важные нюансы старта Брэда Тэнди, включая тестирование измерителя скорости, проведенное на пловцах, прошедших обучение в Университете Индианы.
Ваш в плавании,
Гэри-старший
5 Корректировки тела для улучшения результатов замахов
Многие люди предполагают, что они улучшат свои удары и станут лучшими игроками в гольф на поле, но это не совсем так.На самом деле курс — это то место, где вы видите результат своей практики и улучшений, а не то, где они происходят. Вместо этого, если вы хотите улучшить свой замах, вам придется пойти на тренировочное поле. Практика, которую вы здесь получите, а не время, проведенное на поле, изменит вашу игру в гольф. Диапазон — идеальное время, чтобы немного изменить свою стойку, мощность замах и многое другое. Эти небольшие корректировки могут превратиться в гораздо лучшую игру, если вы будете их практиковать.
Давайте взглянем на пять регулировок, которые помогут добиться желаемого поворота.
Проверьте выравниваниеКак и в случае с автомобилем, вы должны быть уверены, что все выровнено правильно. Если ваши плечи, бедра, ступни и лицо клюшки не выровнены правильно, ваши удары не будут попадать туда, куда вы хотите, или не обладают достаточной силой, чтобы послать мяч на необходимое расстояние. Вы можете использовать палку для выравнивания, чтобы проверить свою стойку, но если у вас ее нет, подойдет все, что имеет прямую кромку. В идеальной стойке, если кто-то смотрит на вас сбоку, он не сможет увидеть вашу заднюю ступню, ногу или плечо.
Подключайтесь к качелямКогда вы размахиваете клюшкой, вы хотите, чтобы ваши руки, кисти, дубинка и ваше тело двигались вместе. Когда вы качаете клюшкой вперед и назад, она должна выглядеть как единое целое. Если вы не связаны таким образом, у вас не будет такого контроля над своим замахом. Это означает, что ваш мяч может отклониться от курса или может не иметь скорости, необходимой для перехода на грин.
Одна из распространенных проблем, с которыми многим приходится работать, — это то, что запястья и руки используются чаще, чем плечи.В идеале вы не должны сильно двигать запястьями, пока булава не будет параллельна земле. Когда он достигнет этой точки, согните запястья и используйте плечи, спину и предплечья, чтобы закончить поднимать и опускать клюшку.
Повернитесь спинойКогда вы отбиваете мяч, вы не просто махаете клюшкой вперед и назад. Вместо этого вы фактически поднимаете его вверх и почти позади своего тела. Чтобы полностью выполнить это, вы не хотите, чтобы ваше тело блокировало движение вашей дубинки к цели.Ваша верхняя часть тела должна сделать полный поворот, когда вы откидываетесь назад. Не полагайтесь только на свои руки, чтобы получить необходимую силу — положитесь на них спиной!
Продолжение движения бедрамиВаши запястья, руки, плечи и спина играют определенную роль в развороте, но при махе вниз играют роль бедра. Когда вы опускаете клюшку, позвольте бедрам вращаться. Для многих это означает, что их правое бедро будет двигаться к мячу, а левое — назад.Это не для всех — некоторые игроки больше сосредотачиваются на одном бедре, чем на другом. Вы хотите определить, что лучше всего подходит для вас.
Даунсвинг похож на бросок в баскетбол или удар по теннисному мячу — вы сделали все, что могли, чтобы настроить его идеально, но как только начинается даунсвинг, вы должны надеяться, что все пойдет по плану, потому что у вас нет большой контроль над этим. Если вы обнаружите, что что-то идет не по плану, возможно, вам придется внести еще несколько корректировок в свой ход.
Нужно больше точности? Вытяните рукуНаконец, если вы обнаружите, что ваши удары не очень точны, вам, возможно, придется поработать над разгибанием правой руки в конце взмаха. Держите руку прямо, когда она касается мяча и касается его, это поможет вам удерживать все на одной линии. Здесь также некоторые игроки в гольф переносят вес на заднюю ногу, толкая бедро. Это предотвращает опережение мяча верхней частью тела, позволяя усилить удары.
Включение этих регулировок в ваши качелиЕсли вы не довольны своим ходом в гольфе, первое, что нужно сделать, — это внимательно оценить, что не так. Возможно, вам понадобится попросить кого-нибудь наблюдать за тем, как вы качаетесь, или даже снимать на видео ваши качели под разными углами, чтобы вы могли внимательно увидеть, что вы делаете и как выровнено ваше тело. Если вы выявляете проблемы, вы можете сосредоточиться на их устранении по очереди, а не пытаться изменить все в своем движении сразу.Это дает вашему телу время для включения изменений в вашу мышечную память и позволяет вам внимательно изучить и усовершенствовать настройку. Однако, как только вы внесете изменения в свой замах, вы обязательно увидите улучшение.
Кинематика поворота плеча рычага в миозине VI
Значение
Миозин VI (MVI), молекулярный мотор, неисправность которого связана с глухотой, движется по актиновой нити, питаемой АТФ. Хемомеханическая трансдукция завершается силовым ударом, при котором моторный домен претерпевает конформационный переход, преувеличенный плечом рычага.Мы выполнили моделирование рабочего хода MVI, показав, что плечо рычага претерпевает почти свободное вращательное распространение, которое лишь слабо смещается остальной частью двигателя. Наша модель дает молекулярную картину рабочего хода MVI, которая количественно согласуется с экспериментами, демонстрирующими податливость плеча рычага. Наши результаты дают представление о широком распределении размера шага MVI.
Abstract
Миозин VI (MVI) — единственный известный член суперсемейства миозинов, который после димеризации процессивно идет к заостренному концу актинового филамента.Передняя головка димера направляет заднюю головку вперед с силовым ходом, конформационное изменение моторной области, преувеличенное плечом рычага. Используя уникальную крупнозернистую модель рабочего хода одного MVI, мы обеспечиваем молекулярную основу его подвижности. Мы показываем, что рабочий ход происходит в два основных этапа. Во-первых, моторная область достигает конформации после инсульта без направления плеча рычага вперед; и, во-вторых, плечо рычага достигает ориентации после удара, претерпевая вращательную диффузию.Из анализа траекторий мы обнаруживаем, что потенциал, который направляет вращающееся плечо рычага к конформации после инсульта, почти плоский, подразумевая, что вращение плеча рычага в основном не связано с двигательной областью. Поскольку обратная нагрузка, сравнимая с наибольшим межголовым натяжением в димере MVI, предотвращает вращение плеча рычага, наша модель предполагает, что плечо рычага с ведущей головкой димера MVI не связано, в соответствии с выводом, сделанным из поляризованного полного внутреннего отражения. флуоресцентные (polTIRF) эксперименты.Без какого-либо настраиваемого параметра наше моделирование приводит к количественному согласию с экспериментами polTIRF, что подтверждает структурные идеи. Наконец, в дополнение к предсказаниям, которые можно проверить, мы также обсуждаем последствия нашей модели для объяснения широкого распределения размеров шага в пошаговом шаблоне MVI.
Как и их аналоги динеины и кинезины, миозины представляют собой молекулярные моторы, которые преобразуют химическую энергию, собранную при гидролизе АТФ, в механическую работу. Они делают это, проходя цикл реакции (рис.1) с участием гидролиза АТФ, связанного со связыванием и отсоединением от нитчатого актина (F-актина) (1, 2). Миозины, содержащие гидролизованный АТФ, связываются с F-актином, высвобождают продукты гидролиза АТФ и претерпевают структурные изменения, известные как силовой удар. Структурное изменение N-концевой части моторного домена, где находятся сайты связывания актина и нуклеотидов, передается в конвертерный домен, который переходит из состояния предпускового удара (PrePS) в состояние пост-силового удара (или строгости, R) конформация.Движение конвертера усиливается большим колебанием плеча рычага, продолговатого домена, связанного с легкими цепями или кальмодулинами (CaMs). Когда новая молекула АТФ связывает безнуклеотидный миозин (в состоянии R), мотор отделяется от актина, и он готов начать новый цикл.
Рис. 1.Реакционный цикл МВИ. Цветовой код: грифельно-синий, моторный домен; красный, ins1; темно-синий, конвертер; оранжевый, ins2- и ins2-связанный CaM; зеленый — домен IQ и CaM, связанный с IQ; черный, актин. Структуры Protein Data Bank (PDB), использованные для создания этого рисунка, — это 4ANJ (41) и 2BKI (33) для моторного домена, плечо рычага MVI было извлечено из 3GN4 (23), а F-актин был заимствован из 1MVW ( 48).Обе модели PrePS и R были сопоставлены с миозином, связанным с актином в 1MVW.
Большая часть работ по немышечным миозинам была сосредоточена на миозине V (MV). Однако после открытия, что миозин VI (MVI) имеет необычную структуру, возрос интерес к подвижности MVI. Помимо своей биофизической важности, MVI участвует в большом количестве клеточных функций у разных организмов (3, 4). Например, MVI участвует в эндоцитозе, сперматогенезе, миграции клеток, организации цитоскелета и аппарата Гольджи, а также в локализации белка.Он также играет роль в поддержании стереоцилий клетки внутреннего уха. Мутации гена myo6 вызывают глухоту у мышей и людей (3). Влияние мутации, вызывающей глухоту, на цикл MVI было недавно выяснено, что позволяет предположить механизм, с помощью которого функционирование мутанта затруднено (5). MVI сверхэкспрессируется в раковых клетках яичников, и ингибирование его экспрессии снижает склонность опухоли к диссеминации (6). MVI также сверхэкспрессируется в раковых клетках простаты (7).
Некоторые миозины выполняют свою физиологическую функцию как мономеры, а другие образуют димеры, которые ходят процессивно, т.е. они делают несколько последовательных шагов на полярном треке F-актина, не отщепляясь. MVI, мономерный в растворе (8), может образовывать димеры (9⇓⇓⇓ – 13), способные к процессуальному движению (9, 10).
Процессивные двигатели демонстрируют высокий коэффициент заполнения (двигатель проводит большую часть своего цикла, тесно связанный с F-актином), стробирование (связь между двумя головками) и рабочий ход (2). Хотя в управлении двигателем с высокой продолжительностью включения может не быть необходимости, это, вероятно, увеличит эффективность двигателя и длину пробега (14).Недавняя крупнозернистая модель MVI предоставила структурные доказательства того, что высвобождение АДФ ограничено (15), что согласуется с экспериментами (16) и кинетическими моделями (14), хотя также было высказано предположение, что блокирование связывания АТФ составляет стробирующий механизм в MVI ( 17).
В дополнение к отображению стробирования и высокой продолжительности включения (17, 18), MVI демонстрирует ряд поразительных особенностей, которые отличают его от других процессивных миозинов, таких как хорошо изученный MV: ( i ) Для начала, тогда как MV и все другие члены суперсемейства миозинов движутся к зазубренному концу (или положительному концу) F-actin, MVI шагает к заостренному концу (или отрицательному концу) (2, 19).( ii ) Архитектура плеча рычага значительно различается между MV и MVI (20). В MV имеется шесть CaMs или IQ-доменов, связанных с легкой цепью, которые составляют плечо рычага, длина которых соизмерима с расстоянием 36 нм в повторе F-actin. MVI имеет только один домен IQ, но на шаге покрывает то же расстояние, что и MV, предполагая, что другие элементы вносят вклад в плечо рычага. Хотя их происхождение остается неясным (20–26), недавние исследования функции MVI подтверждают важность проксимального хвоста как удлинения плеча рычага in vivo (27).( iii ) Кроме того, распределение размеров шага у MV дикого типа намного уже, чем у MVI (9, 10, 20). ( iv ) И, наконец, и MV, и MVI перемещаются процессивно за счет ручного механизма (28⇓ – 30), но в MVI также имеются свидетельства наличия ступеней, похожих на дюймовые червяки (31).
Большинство нетрадиционных механических свойств MVI приписываются вставке 1 и вставке 2 (ins1 и ins2) (рис. 2), двум уникальным фрагментам в суперсемействе миозинов (19). Одномолекулярные эксперименты (32) и кристаллические структуры состояний R (33) и PrePS (34) указали ins2 как ключевой структурный элемент, ответственный за изменение направленности MVI во время шага.Действительно, ins2 оборачивается вокруг домена преобразователя, эффективно поворачивая направление качания назад, а его удаление изменяет направленность движения MVI. Кинетические эксперименты показывают, что ins1 играет ключевую роль в определении высокой продолжительности включения MVI и стробирования (17, 35).
Рис. 2.Последовательность и структурные модели МВИ. ( A ) Последовательность смоделированного MVI со связанным CaM. ( B и C ) Модели PrePS ( B ) и R ( C ), используемые в симуляциях.Цветовой код такой же, как на рис. 1. Общее количество остатков в белке составляет 834. Каждый CaM имеет 145 остатков. Общее количество остатков для наших моделей PrePS и R составляет 1124. F-актин не был частью модели, но он показан здесь для справки. Красные сферы показывают остатки N785 и K834, начало и конец плеча рычага в нашей модели. Серые и черные сферы показывают P66 и A73 IQ-связанного CaM. Размер четырех сфер увеличен для улучшения видимости. В исх.ось и ориентирована в отрицательном направлении, смотря на заостренный конец нити.
Некоторые существенные особенности шагового механизма MVI до конца не изучены, в том числе необычно большое распределение размера шага и чередование шагов с ручным управлением и шагами «дюймового червя». Хотя модели димера, включающие гибкость плеча рычага MVI, способны воспроизводить известные особенности подвижности MVI (36), эксперименты с использованием химерного MVI с плечом рычага MV показали, что распределение размеров ступенек было аналогично дикому типу ( 32, 37), предполагая, что неуловимая структура плеча рычага MVI не может быть ключевым ингредиентом, необходимым для объяснения своеобразного шагового механизма MVI.После экспериментальных исследований, структурного моделирования и моделирования можно сделать вывод, что отсоединение плеча рычага ведущей головки MVI от движения преобразователя или податливость плеча рычага, а также конформационный переход в преобразователе могут способствовать к распределению больших ступеней (30, 38⇓⇓ – 41). Более того, согласно недавно предложенной модели (31, 37), короткие шаги происходят, если свободная задняя головка димера миозина связывает F-actin, в то время как плечо рычага связанной с актином ведущей головки остается в ориентации до удара.И наоборот, поворот плеча рычага со связанной головкой в положение R приводит к большим шагам.
Хотя экспериментальные данные подтверждают модель разъединения или податливости плеча рычага (30, 38⇓⇓ – 41), прямые доказательства требуют структурной модели, способной описывать динамику поворота плеча рычага. Мы используем комбинацию крупномасштабного (CG) моделирования и теории, чтобы исследовать динамику поворота плеча рычага в MVI. В последние годы многочисленные симуляции на различных системах показали надежность компьютерных моделей в выявлении основных особенностей динамики макромолекул (42–47).Мы подготовили модель для PrePS и R конформации, которая включает плечо рычага до ins2 и домен IQ (рис. 2 B и C ). Мы смоделировали силовой удар, индуцируя переход от PrePS к конформации R, игнорируя любую промежуточную конфигурацию и F-актин (см. Материалы и методы для более полного обсуждения). Мы сгенерировали 96 траекторий перехода PrePS → R для изучения рабочего хода — для обсуждения сходимости результатов см. SI Приложение , раздел 4 и SI Приложение , Рис.S14. Чтобы понять влияние обратной нагрузки на вращение плеча рычага, мы сгенерировали 96 траекторий перехода PrePS → R с плечом рычага, подверженным действию силы сопротивления 6 пН. Наши результаты показывают, что переход PrePS → R происходит в два основных этапа: на первом этапе двигательный домен достигает состояния R, в то время как плечо рычага разъединено. На втором этапе плечо рычага диффундирует к конфигурации R-состояния с небольшим руководством со стороны моторной области. Наша модель приводит нас к предположению, что ступени, похожие на дюймового червяка, могут происходить, когда свободная головка связывает F-актин до того, как несвязанное плечо рычага поворачивается в конформацию R-состояния, которая определяет прямую связь между короткими (или похожими на дюймового червя) ступенями и отсоединение плеча рычага.Результаты нашего моделирования не только выгодно отличаются от экспериментов, но и дают точные, проверяемые прогнозы.
Результаты
Вращение рычага происходит в два основных этапа.
Мы отслеживаем динамику перехода PrePS → R, используя функцию структурного перекрытия (χ) по отношению к состоянию R, задаваемую формулой (49) χ (t) = 2NB2−5NB + 6∑i = 1NB − 3∑j = i + 3NBΘ (| rij (r) −rij (R) | −a), [1] где NB — количество гранул в модели CG (за исключением тех, которые не были разрешены в кристаллической структуре, см. приложение SI , сечение 1), rij (t) — расстояние между бусинами i и j в момент времени t, rij (R) — расстояние в состоянии R, Θ — функция Хевисайда, а a = 2Å — допуск.Суммирование в уравнении. 1 — это пары бусинок, которые находятся по крайней мере на три остатка друг от друга в последовательности (общее количество таких пар является обратной величиной префактора в уравнении 1 ). В состоянии PrePS χ≈0,62, а в состоянии R χ≈0,34. Временной ход χ показывает, что переход PrePS → R происходит в два этапа (черная линия на рис. 3 A ). В течение нескольких микросекунд χ уменьшается до ≈0,55 (рис. 3 A , вставка ) и колеблется около этого значения в течение длительного времени, пока не подвергнется еще одному быстрому переходу, ведущему в состояние R.Функция структурного перекрытия колеблется около 0,34 до конца моделирования. Подобные закономерности обнаруживаются на всех траекториях.
Рис. 3.Динамика перехода PrePS → R. ( A ) Зависимость χ от времени показана черным цветом. Красная стрелка указывает конец первого перехода, а зеленая и синяя стрелки соответствуют началу и концу второго перехода соответственно. A также показывает временные кривые расстояния между C63 и R708 (красные квадраты), T754 и D24 связанного с ins2 CaM (зеленые кружки) и V140 и D58 CaM, связанного с ins2 (синие треугольники). A , Вставка показывает начальный переход по χ. ( B , Left ) Гистограмма τχ1. ( B , Центр ) Гистограмма τχ2 − τχ1. ( B , Правый ) Гистограмма τχ3 − τχ2. Здесь τχ1 — конец первого перехода, а τχ2 и τχ3 — соответственно начало и конец второго перехода, контролируемого χ.
Обозначим время завершения первого шага как τχ1, время до начала второго шага как τχ2, а время завершения второго шага как τχ3.На практике τχ1 (τχ3) измеряется, когда χ впервые достигает 0,55 (0,34) (красная и синяя стрелки на рис. 3 A ). Время начала второго шага, τχ2, измеряется как последний раз, когда траектория пересекает 0,55 перед достижением 0,34 (зеленая стрелка на рис. 3 A ). Гистограммы τχ1, τχ2 − τχ1 и τχ3 − τχ2 показаны на рис. 3 B . Обратите внимание на разницу в масштабе абсцисс. Ясно, что первый переход происходит быстро (среднее значение τχ1 равно τχ1⟩ = 2,6 мкс, при SD στχ1 = 1.2 мкс), за которым следует долгое время ожидания (⟨τχ2 − τχ1⟩ = 178 мкс, στχ2 − τχ1 = 122 мкс) перед еще одним быстрым переходом (⟨τχ3 − τχ2⟩ = 4,9 мкс, στχ3 − τχ2 = 2,4 мкс). Происхождение большого распределения τχ2 − τχ1 обсуждается в следующем разделе.
Структурные переходы на двух этапах.
Чтобы охарактеризовать структурное происхождение двух ступеней, наблюдаемых на временной шкале χ (t) во время перехода PrePS → R, мы отслеживаем следующие три расстояния между бусинами, которые образуют контакты в состоянии R, но не в состоянии PrePS. : ( i ) dRED — это расстояние между C63 и R708 (красные сферы на рис.4), ( ii ) dGREEN — это расстояние между T754 и D24 ins2-связанного CaM (зеленые сферы на рис. 4), а ( iii ) dBLUE — это расстояние между V140 и D58 границы ins2. CaM (синие сферы на рис. 4). Когда преобразователь переходит из PrePS в конформацию R-состояния, образуется контакт dRED. Образование контакта dGREEN указывает на начальное замыкание плеча рычага на область двигателя, которое завершается, когда формируется контакт dBLUE. Временные характеристики этих трех расстояний для конкретной траектории показаны на рис.3 A , из которого следует, что формирование контакта dRED совпадает с первым шагом в уменьшении χ (t), а второй этап χ (t) происходит примерно в то же время, когда образуются dGREEN и dBLUE. Чтобы определить, справедливо ли это наблюдение для всех траекторий, мы извлекаем из каждой траектории время устойчивого образования трех контактов. Под стабильным мы подразумеваем, что как только контакт сформирован, он остается неповрежденным с небольшими колебаниями (на практике мы контролируем стабильные контакты, проверяя первый момент формирования контакта, так что в остальной части моделирования контакт остается ближе к контакту. Идеальное расстояние в состоянии R, чем самое большое расстояние, исследованное во время перехода PrePS → R).Мы называем время стабильного образования dRED, dGREEN и dBLUE как t1, t2 и t3, соответственно, и исследуем корреляции между t1 и τχ1, t2 и τχ2, а также t3 и τχ3. Рис. 5 A показывает, что существует хорошая корреляция между t1 и τχ1, за исключением нескольких сплошных кружков, которые представляют все те траектории, на которых плечо рычага начинает замыкаться на область двигателя до вращения области преобразователя. происходит, то есть до образования dRED. В этих случаях начальное падение χ (t) отражает образование контакта dGREEN.В трех из пяти случаев (синие сплошные кружки на рис. 5 A ) первоначальный контакт dGREEN является временным, что означает, что он быстро разрывается, преобразователь вращается и, наконец, восстанавливается; таким образом, в конечном итоге устойчивое образование контакта dRED предшествует контакту dGREEN. В двух случаях, показанных черными сплошными кружками на рис. 5 A , контакт dGREEN устойчиво образуется до dRED, и он не ломается. Коэффициент корреляции Пирсона и линейная аппроксимация, выполненная в отсутствие закрашенных кружков, показывает, что в большинстве случаев (91/96 траекторий) dRED формируется около τχ1.Это говорит о том, что при t≈τχ1 преобразователь находится на стороне плюса области двигателя MVI (рис. 2 B и C ). Тщательное изучение кристаллических структур до и после перехода PrePS → R (33, 34) показывает, что конвертер претерпевает вращение, а также подвержен конформационным изменениям с так называемой P-складки на R-складку. Расстояние dRED может отслеживать только перемещение преобразователя, но мы показываем в приложении SI , разделы 2 и 3 и в приложении SI , рис.S6 – S8, что около τχ1 преобразователь и моторный домен претерпевают необходимое вращение и структурный переход для достижения конформации R-состояния. Мы пришли к выводу, что в большинстве случаев первый шаг поворота плеча рычага соответствует перемещению преобразователя из PrePS в R-конфигурацию.
Рис. 4.Мониторинг ключевых структурных изменений при переходе PrePS → R. Расположение атомов Cα у C63 и R708 показано красными сферами, а расположение атомов T754 и D24 ins2-связанного CaM показано зелеными сферами.Синие сферы показывают положения Cα V140 и D58 ins2-связанного CaM, соответственно. Пунктирные линии соединяют бусины, входящие в контакты dRED, dGREEN и dBLUE. Для улучшения видимости расположение сфер также выделяется меткой R, G или B. A показывает состояние PrePS, а B показывает состояние R.
Рис. 5.Корреляция между переходом по χ и структурными изменениями MVI. ( A ) График зависимости τχ1 от t1. Синие (черные) сплошные кружки обозначают результаты моделирования, в которых временно (постоянно) контакт dGREEN образуется раньше контакта dRED.Наклон линейной аппроксимации составляет ≈0,78, в то время как точка пересечения составляет ≈0,86 мкс. ( B ) График зависимости τχ2 от t2. Наклон линейной аппроксимации составляет ≈1,00, а точка пересечения составляет ≈2,6 мкс. ( C ) τχ3 в зависимости от t3. Наклон линейной аппроксимации составляет ≈1,00, а точка пересечения составляет ≈0 мкс. Линейные аппроксимации и коэффициенты корреляции вычисляются без синих сплошных кружков в A и со всеми кружками в B и C .
Пирсоновские корреляции t2 с τχ2 и t3 с τχ3 показаны на рис.5 B и C . Два коэффициента корреляции очень близки к единице, и линейная аппроксимация дает небольшие пересечения (легенда рис. 5). Следовательно, вокруг τχ2 образуется контакт dGREEN, за которым следует контакт dBLUE при t≈τχ3.
Анализ, проведенный до сих пор, отслеживая χ и формирование контактов dRED, dGREEN и dBLUE, предполагает, что мы можем разделить наши траектории на два класса: ( i ) χ соответствует перемещению домена преобразователя из PrePS в положение R (образование контакта dRED).В редких случаях (3/96) мы наблюдали, что движению преобразователя предшествует временное образование контакта в R-состоянии между плечом рычага и моторным доменом (контакт dGREEN). После перемещения преобразователя после долгого времени ожидания ins2 замыкается на область двигателя, чтобы сформировать состояние R (стабильное образование контактов dGREEN и dBLUE). Следовательно, первый этап — это переход рабочего хода двигателя (включая преобразователь), а во время второго этапа плечо рычага достигает конформации R-состояния.Это говорит о том, что на всех этих траекториях (94/96) движение плеча рычага «не связано» с переходом PrePS → R преобразователя. Мы называем эти траектории несвязанными. Примеры несвязанных траекторий показаны в фильмах S1 и S2 и приложении SI, рис. S11 и S18. ( ii ) В меньшинстве траекторий (2/96) устойчивый контакт, подобный R-состоянию, между ins2 и доменом двигателя формируется перед переключением положения преобразователя. Эти траектории классифицируются как «связанные», потому что в этом механизме взаимодействие между плечом рычага и областью двигателя приводит к быстрому повороту плеча рычага, направляемому движением преобразователя.Фильмы S3 и S4 и SI Приложение , рис. S12 и S19 показывают примеры связанных траекторий.
Число связанных траекторий может быть искусственно увеличено путем изменения потенциала, который вызывает переход PrePS → R ( SI Приложение , раздел 1 и SI Приложение , рис. S2 – S4). Однако конформация плеча рычага, образующегося на связанных траекториях, не воспроизводила экспериментальные данные [5]. 38 ( SI Приложение , раздел 1 и SI Приложение , Рис.ось и ориентирована в отрицательном направлении. Вид наблюдателя, использованный в качестве эталона, описан на рис. 2 B и C . Чтобы следить за движением плеча рычага, мы отслеживаем положение N785 (в ins2, показано красным на рис. 2 B и C ) и K834 (последние остатки IQ-домена показаны красным валик на рис.2 B и C ). Примеры траекторий ( SI Приложение , рис. S9, S11 и S18 и видеоролики S1 и S2) показывают, что N785 перемещается в течение t <τχ1 из состояния PrePS почти в конфигурацию R-состояния.Когда t превышает τχ1, он быстро находит положение равновесия R-состояния, колеблясь около этого значения до конца моделирования. С другой стороны, K834 совершает большое вращательное движение вокруг преобразователя от значения в состоянии PrePS до значения в состоянии R ( SI, приложение , рисунки S9, S11 и S18 и фильмы S1 и S2). Мы пришли к выводу, что после начального шага в переходе PrePS → R, N785 служит шарниром, вокруг которого вращаются дистальная часть ins2 и домен IQ, пока не достигнут состояния R.Чтобы отслеживать вращение плеча рычага наиболее интуитивно, мы описываем вектор, соединяющий N785 и K834, в сферических координатах. Поскольку расстояние между N785 и K834 примерно постоянно во время перехода PrePS → R ( SI Приложение , рис. S10, Top ), мы описываем движение в состояние R несвязанного плеча рычага как вращение жесткого стержня с изменения высотного угла θ и азимутального угла ϕ (рис. 6 A ). Поскольку соответствующее движение плеча рычага происходит в направлении F-актина, мы выбираем θ в качестве параметра порядка и описываем динамику плеча рычага на единичной окружности.
Рис. 6.Извлечение потенциала, приводящего во вращение. ( A ) Определение углов θ и ϕ. Декартовы оси показаны красным, зеленым и синим цветом. Направление плеча рычага показано жирной красной линией, а положение N785 и K834 показано сферами. Обратите внимание на рис. 2 B и C , что ось z параллельна F-actin, поэтому угол θ указывает угол между плечом рычага и актиновой нитью. Источник и сток модели Фоккера – Планка (FP) приблизительно показаны фиолетовым и оранжевым цветом соответственно.( B ) Вероятность ρr (θ). Результат компьютерной симуляции показан синими сплошными кружками. Подгонка данных, полученных с использованием приложения SI, приложение , уравнение. S11 , красного цвета. Серыми жирными линиями показаны значения θi = 2,25 и θf = 0,25. ( C ) Полученная свободная энергия соответствует ρr (θ). Серые толстые линии такие же, как у B .
Энергетика поворота рычага.
Вращающееся плечо рычага подвергается воздействию свободной энергии F (θ), которая управляет его движением.Мы извлекаем эту свободную энергию из траекторий, чтобы определить степень связи между плечом рычага и двигательной областью. Предполагая, что в начале вращения плечо рычага находится под углом θ = θi, а в конце высотный угол равен θ = θf, и игнорируя роль азимутального угла, большое (F (θi) −F (θf ) >> kBT), свободная энергия спуска предполагает, что качели управляются. Напротив, если свободная энергия плоская (F (θi) -F (θf) ≈kBT), то плечо рычага разъединяется в направлении θ.Чтобы извлечь свободную энергию F (θ), мы не можем просто вычислить логарифм равновесного распределения вероятностей ρ (θ), потому что как только плечо рычага достигает состояния R, мы не наблюдаем переходов в состояние PrePS. Следовательно, система вышла из равновесия, и нам нужно принять другую стратегию для извлечения F (θ). Мы принимаем процедуру, изложенную в исх. 50 (подробности в SI Приложение , раздел 4). Мы генерируем стационарное распределение вероятностей на единичной окружности ρr (θ) из смоделированных переходов PrePS → R.Траектории инжектируются под углом θi = 2,25 рад (близко к состоянию PrePS, θPrePS≈2,55 рад) и удаляются, когда они пересекают θf = 0,25 рад (близко к состоянию R, θR≈0,04 рад). Мы предполагаем, что стационарная вероятность ρr (θ) является решением уравнения Фоккера – Планка (FP) с соответствующими граничными условиями для учета впрыска и удаления траекторий (50). Поскольку мы знаем ρr (θ) из моделирования, мы получаем свободную энергию F (θ), решая уравнение FP. Чтобы решить уравнение FP, мы предполагаем, что коэффициент диффузии D является постоянным.На рис. 6 B показано распределение вероятностей ρrCG (θ), полученное в результате анализа компьютерных симуляций (синие сплошные кружки). Как подробно описано в Приложении SI , раздел 4, мы аппроксимируем свободную энергию F (θ), чтобы получить распределение вероятностей ρr (θ) (рис. 6 B , красная линия), которое точно воспроизводит результаты моделирования.
Профиль F (θ) показывает, что от θ = 2,25 рад (≈129o) до θ≈π / 4 (≈45o) большая часть качания происходит на почти плоском профиле свободной энергии (рис.6 С ). Для θ <π / 4 имеется неглубокий минимум ≈1 ккал / моль, который приводит в движение последнюю часть колебания. Мы заключаем, что качание происходит без сильного взаимодействия с двигательной областью, подразумевая, что движение плеча рычага в основном не связано, что качественно проявляется в траекториях ( SI Приложение , рис. S11 и S18 и фильмы S1 и S2). Это означает, что большую часть поворота плечо рычага вращается стохастически, сохраняя шарнир вокруг N785.Захват к состоянию R происходит только тогда, когда плечо рычага достаточно близко к двигательной области. Стохастичность динамики объясняет большую ширину распределения τχ2 − τχ1.
Чтобы извлечь постоянную диффузии, мы сравниваем смоделированное среднее время первого прохождения (MFPT) τrCG (θ) и время, возникающее в результате диффузии псевдочастицы в потенциале F (θ). Теоретическая MFPT получается путем решения одномерного уравнения FP с использованием постоянного коэффициента диффузии (51), τFP (θ) = D − 1∫θfθdθ′eβF (θ ′) ∫θ′πdθ ″ e − βF (θ ″) .Подгоняя D в τFP (θ) к τrCG (θ), мы получаем D≈0,011 / мкс ( SI Приложение , раздел 4). Хорошее соответствие между τFP (θ) и τrCG (θ), указанное в Приложении SI , рис. S13, предполагает, что разумно предположить, что D является постоянным.
Хотя мы построили одномерную модель, которая отражает особенности компьютерной симуляции, мы подчеркиваем, что вращение происходит на сфере, что было бы лучше зафиксировано в 2D-модели. Действительно, азимутальная составляющая может играть важную роль во вращении плеча рычага.
Сравнение с экспериментами.
Мы используем наше моделирование для сравнения с экспериментами, исследующими ступенчатый механизм димеров MVI (38). Чтобы следить за ориентацией плеча рычага во время шагания, CaM, связанный с IQ доменом MVI, метили бифункциональным родаминовым (BR) зондом (38). Точки крепления — P66 и A73, они показаны на рис. 2 B и C черным и серым цветом соответственно. Sun et al. (38) вывели динамику вращения плеча рычага из ориентационных изменений BR, используя поляризованную флуоресценцию полного внутреннего отражения (polTIRF).Это дало углы θ (вдоль направления актина, обозначенного β в ссылке 38) и угол ϕ (азимутальный угол обозначен как α в ссылке 38). [Обратите внимание, что θ и ϕ — это углы зонда BR относительно референсной системы актина (рис. 2 B и C ). В исх. 38 греческие буквы θ и ϕ используются для обозначения угла плеча рычага в системе отсчета актина, а α и β используются для датчика. Мы сравниваем здесь с α и β из исх. 38]. Были идентифицированы головки димеров в ведущей и задней позициях и извлечены соответствующие распределения углов θ и ϕ.Согласно кинетическим экспериментам (52), как задняя головка (TH), так и ведущая головка (LH) находятся в состоянии, связанном с ADP. В TH поворот плеча рычага уже произошел, и он направлен к заостренному концу F-актина. Напротив, ЛГ, удерживаемый под напряжением связанным с актином TH, находится в несвязанном (U) состоянии.
Чтобы сравнить наше моделирование с экспериментами, мы воспроизвели измерения, отслеживая ориентацию единичного вектора, соединяющего A73 и P66 (рис. 2 B и C ).Углы θ и ϕ зонда BR в кристаллической структуре с R-состоянием различаются между расчетной моделью (θRCG и ϕRCG) и значениями, использованными в исх. 38 (θREXP и ϕREXP). Два фактора, вероятно, способствуют этому несоответствию: ( i ) Выравнивание с F-актином и выбор системы сравнения не идентичны в экспериментах и моделированиях (наши оси повернуты по сравнению с осями в ссылке 38, поэтому ϕ≈α + π / 2) и ( ii ) мы извлекли углы из относительного положения α-углеродов P66 и A73 из CaM, тогда как эксперименты, по всей вероятности, обнаруживают ориентацию β-углеродов (53) .
Чтобы описать TH, мы рассматриваем конформации R-состояния, выбранные в моделировании только при t> τχ3, чтобы гарантировать, что состояние R достигнуто. Чтобы получить образец U-состояния ведущей головки, мы выполнили 96 симуляций поворота плеча рычага, приложив к кончику плеча рычага (K834) обратную нагрузку в 6 пН (подробности см. В приложении SI, приложение , раздел 1). , имитирующий эффект межголового натяжения. Механическая сила (fL), принятая для описания состояния U, противодействует вращению плеча рычага, прикладывая импульс силы величиной ≈rLAfL, где мы рассматриваем плечо рычага rLA≈7.5 нм ( SI Приложение , Рис. S10, Top ). Если мы рассмотрим все плечо рычага MVI (rLA≈18 нм), сила, генерирующая такой же резистивный момент, равна fL≈2,5 пН, что близко к верхней границе межголового натяжения, оцененной экспериментально [fL≈2,2 пН (16 )]. Это говорит о том, что величина нашей обратной нагрузки не является нереально высокой. Мы показываем в приложении SI , разделы 2 и 3, что обратная нагрузка 6 пН не влияет на первую ступень перехода PrePS → R (приложение SI , рис.S6 – S8 и S20, приложение SI , раздел 7 и видеоролики S5 и S6), но этого достаточно, чтобы предотвратить возникновение прямого вращения плеча рычага в 95 из 96 случаев ( SI Приложение , раздел 1 и SI Приложение , рис. S3). Таким образом, мы сгенерировали ансамбль U-образных конфигураций как набор структур из моделирования, проведенного с обратной нагрузкой 6 пН, выбрав только конфигурации для t> τχ1, для которых вращение плеча рычага не было завершено.
Сравнение экспериментальных данных и моделирования представлено на рис. 7. Мы сравнили следующие величины, указанные в ссылке. 38: ( i ) распределение вероятностей для LH и TH угла θ (рис. 7 A ), ( ii ) распределение вероятностей для изменения угла ϕ после одного шага (Δϕ, рис. .7 B ) и ( iii и iv ) распределение вероятностей для изменения после двух шагов угла θ ( iii ) (Δ2θ, рис.7 C ) и угол ϕ ( iv ) (Δ2ϕ, рис. 7 D ). Поскольку в экспериментах нет контроля азимутального угла посадки MVI на актине, экспериментальное распределение угла ϕ для LH и TH почти плоское на 180o. При моделировании мы всегда начинаем с MVI, параллельного оси x ; таким образом, сравнение рассчитанных и измеренных распределений может ввести в заблуждение ( SI Приложение , раздел 5 и SI Приложение , Рис.S15).
Рис. 7.Сравнение результатов моделирования и экспериментов. В A – D гистограммы показывают данные, извлеченные из рисунков 3 и 4 исх. 38, а линии показывают результаты наших симуляций и анализов. Красные и синие линии получены непосредственно из моделирования, а черные линии — из нашей модели прямой ступени димера MVI ( SI Приложение , раздел 5 и SI Приложение , уравнения S17 и S18 ), на основе подгонки смоделированных распределений в A и в приложении SI , рис.S15. ( A ) Распределение θ (β в ссылке 38) задней (красной) и передней (синей) головок. Данные моделирования были подогнаны с использованием нормальных распределений. Показаны значения средних и дисперсий. Среднее значение распределения обозначено чертой, а стандартное отклонение обозначено как σ. ( B ) Распределение изменения ϕ после одного шага ( SI Приложение , уравнение S17 ). ( C ) Распределение изменения θ после двух шагов ( SI Приложение , уравнение. S18 ). ( D ) Распределение изменения ϕ после двух шагов ( SI Приложение , уравнение S18 ).
Для сравнения и мы напрямую извлекли распределения вероятностей из компьютерных симуляций состояния U и R, чтобы описать LH и TH, соответственно. Мы не моделировали последовательные шаги. Таким образом, для описанных выше сравнений ii – iv мы создали простую модель изменений углов после одного или двух шагов, используя два допущения (подробности см. В приложении SI , раздел 5): ( i ) Димер всегда шагов с помощью ручного механизма, и ( ii ) значения углов после шага не зависят от значений до шага.
Мы обнаружили замечательное соответствие между углами θ, полученными при моделировании и эксперименте, как для LH, так и для TH (рис. 7 A ). Наша модель CG четко воспроизводит ориентацию и колебания зонда BR в LH и TH, что подтверждает модель. Следует подчеркнуть, что мы не настраивали какой-либо параметр в модели ЦТ для получения согласия с экспериментами.
Хотя рассчитанные изменения угла ϕ после одного шага согласуются с экспериментальными данными (рис.7 B ), существует разница в интерпретации результатов, основанных на моделировании и экспериментах. Согласно экспериментам, в обоих переходах LH → TH и TH → LH Δϕ может быть положительным или отрицательным с примерно равной вероятностью (таблица 1 в ссылке 38). Из моделирования мы находим, что Δϕ в большинстве случаев (≈97% времени) является положительным (отрицательным) при переходах R → U (U → R). Таким образом, при моделировании в каждый пик P (Δϕ) вносят вклад переходы R → U или U → R, тогда как в экспериментах каждый переход вносит вклад в два пика почти одинаково.Более того, в нашей ступенчатой модели мы предполагаем, что направление межголового натяжения всегда совпадает с F-actin, что подразумевает, что происходит только ступенька 36 нм. Но измеренное широкое распределение размера шага в MVI предполагает, что это не всегда так.
Сравнение распределения вероятностей после двух шагов дает точные результаты для угла θ (рис. 7 C ), тогда как наша модель распределения Δ2ϕ дает несколько более пиковое распределение (рис. 7 D ).Это говорит о том, что в нашей модели пошагового выполнения MVI далеко не так «волнистый», как предполагалось из экспериментов (38). Для более количественного сравнения с экспериментами требуется CG-модель димера MVI в комплексе с F-актином.
Обсуждение
Положение шарнира поворотного механизма.
Мы предполагаем, что положение шарнира, вокруг которого вращается плечо рычага, близко к остатку N785 (расположенному в ins2), в начале области, идентифицированной как гибкая в состоянии PrePS (41).В течение нескольких микросекунд от начала перехода PrePS → R N785 связывает преобразователь, а затем колеблется около среднего положения ( SI Приложение , рис. S9 и S18 – S20). Эксперименты показали, что химерные конструкции, в которых ins2 был усечен до (после) N785 и заменен искусственным плечом рычага, представляют собой двигатели с положительным (отрицательным) концом (14, 54). Таким образом, с точки зрения конструкции расположение петли физически разумно.
Отсоединение рычага во время рабочего хода присуще головке двигателя.
Двухступенчатый переход PrePS → R с разъединением плеча рычага при нулевой обратной нагрузке предполагает, что не наличие связанного с актином TH вызывает разъединение плеча рычага. Наше моделирование показывает, что существует возможность возникновения сопряженного колебания. Однако это маловероятно при нулевой силе и наблюдалось только один раз, когда обратная нагрузка 6 пН была приложена к кончику плеча рычага. Следовательно, механизм разъединения присущ силе такта MVI, и он, вероятно, не связан с закрытием или обратным натяжением.Гибкость плеча левого рычага приписывалась податливости области между плечом рычага и преобразователем (и, возможно, удлинением плеча рычага) (38) или отсоединением плеча рычага от области двигателя (30, 39, 41). В этой статье мы используем «разъединение», потому что мы наблюдаем, что на большинстве траекторий вращение плеча рычага не синхронно с рабочим ходом. С другой стороны, мы наблюдаем, что часть ins2 (примерно до N785) плотно прикреплена к преобразователю во время качания ( SI Приложение , рис.S8, S18 и S20 и фильмы S1, S2, S5 и S6), предполагая, что начальная часть ins2 не отделена от моторного домена. Таким образом, наши симуляции согласуются с изображением гибкого плеча рычага вокруг ins2.
Двухступенчатый механизм.
Было высказано предположение, что рабочий ход MVI происходит с помощью двухступенчатого механизма, в котором сначала вращается преобразователь, а затем, как только обратная нагрузка от связанной с актином задней головки снимается, плечо рычага вращается в направлении Конфигурация R-состояния (41).На этом рисунке утверждалось, что в конце первого шага домен преобразователя находится в конфигурации, подобной состоянию PrePS, но он переместился в положение R-состояния в области двигателя. Мы обнаружили, что домен преобразователя не только перемещается и поворачивается в положение R-состояния в области двигателя ( SI Приложение , раздел 3 и SI Приложение , Рис. S7), но также принимает конформацию, наблюдаемую в R- состояние кристаллической структуры (так называемая R-складка преобразователя; SI Приложение , рис.S7 и S8). Гибкость шарнира позволяет этим изменениям происходить даже при наличии большой обратной нагрузки (приложение SI , рис. S7 и S8 и приложение SI , раздел 3). Стоит отметить, что мы не рассматриваем эффекты азимутальной составляющей обратной нагрузки, которые могут усложнить картину, как мы предложили при сравнении результатов моделирования и экспериментов. Обратите внимание, что данные экспериментов с одной молекулой MVI (8) и крио-ЭМ изображений MV (55) предполагают, что поворот плеча рычага состоит из двух подэтапов: большого перехода к конформации, связанной с АДФ, запускаемой высвобождением Pi, и меньшего шаг, происходящий после выпуска ADP.Наш двухступенчатый механизм игнорирует промежуточное соединение, связанное с ADP, и, таким образом, эти два подэтапа, предложенные экспериментами, включены в наш второй переход, происходящий между τχ2 и τχ3. Нельзя исключить, что конвертерный домен в постинсультном состоянии занимает конформацию P-складки (41). Мы полагаем, что R-кратный преобразователь после инсульта дает более сжатую картину рабочего хода MVI. Для решения этой тонкой проблемы необходимы дополнительные симуляции и эксперименты.
Рычаг MVI вращается стохастически, предпочтительно в сторону F-актина.
Как связанные, так и несвязанные траектории предполагают, что структура конвертерного домена и переход PrePS → R благоприятствуют повороту плеча рычага MVI на стороне актиновой нити, то есть с правой стороны, как видно наблюдателю. на рис. 2 B и C (фильмы S1 – S4 и SI, приложение , рис. S11, S12, S18 и S19). Как уже указывалось в исх. 38, это может придавать MVI экспериментально наблюдаемое правостороннее вращение вокруг актинового филамента (20, 38).В несвязанных симуляциях вращение является стохастическим: азимутальная составляющая плеча рычага претерпевает большие изменения, а плоский ландшафт свободной энергии, исследуемый с помощью компонента θ, предполагает, что вращательная диффузия почти свободна для большей части поворота плеча рычага.
Связь между U-состоянием и появлением коротких (похожих на червяков) шагов.
Модель была предложена для объяснения наличия коротких шагов, похожих на дюймовых червей, в моторике MVI (31). Авт. Предполагают, что короткий шаг происходит, если свободная головка MVI связывает актин, в то время как плечо рычага ведущей, связанной с актином головки все еще находится в PrePS ориентации.Если плечо рычага MVI, связанного с актином, поворачивается в положение R-состояния, димер делает длинный шаг. Эта схема обеспечивает структурную картину, которая охватывает как длинные, так и короткие шаги MVI, что объясняет широкое распределение шагов по размеру. Основываясь на двухступенчатом механизме рабочего хода MVI, мы предполагаем, что короткие (похожие на дюймового червя) шаги происходят, если TH повторно связывает F-actin перед вращением несвязанного плеча рычага, то есть до завершения второго шаг перехода PrePS → R.Если вторая стадия перехода завершена, то димер делает большой шаг. Поскольку гидролиз АТФ и ход восстановления происходят до повторного связывания свободной головки с F-актином, для того, чтобы наше предположение было справедливым, вращение несвязанного плеча рычага должно происходить в масштабе времени, сравнимом со скоростью гидролиза АТФ, которая была оценена как быть ≈10−17s − 1 (56). Временной масштаб перехода из состояния U в состояние R составляет τχ3 − τχ1⟩≈183 мкс, что получается для плеча рычага около 7,5 нм.Большие шаги димеров MVI требуют комбинации традиционного плеча рычага и удлинения плеча рычага примерно на 36 нм для димера (без учета шарнира между двумя плечами рычага и свободной головкой миозина). Если мы предположим, что шкала времени вращения зависит от длины вращающегося стержня как L3 / ln (L / 2a) (57), где a — толщина стержня (с учетом CaM она составляет ∼3 нм), то итоговая шкала времени становится около 2,5 мс, что всего в ≈20 раз быстрее, чем наивысшая граница измеренной скорости гидролиза АТФ.Это предполагает, что вполне вероятно, что TH может связываться до того, как плечо рычага ведущей головки достигнет состояния R (то есть после τχ1, но до τχ3).
Отсоединение рычага и выброса фосфата.
Предполагается, что после отпускания Pi и завершения рабочего хода путь для выхода Pi закрывается, что позволяет избежать повторного связывания Pi и сделать переход необратимым (2). Интересно, что недавняя крио-ЭМ структура MV, связанного с F-actin, указывает на то, что сайт связывания нуклеотидов в постинсультной, ADP-связанной конформации не может связывать Pi даже в отсутствие обратной нагрузки (55).При больших усилиях рабочий ход MV кажется обратным (58). Поскольку структурные данные свидетельствуют о том, что преобразователь механически связан с сайтом связывания нуклеотидов через релейную спираль, индуцированное нагрузкой обращение силового удара будет способствовать восстановлению конформации моторного домена, способного связывать Pi и более легко отделяться от F -актин, что объясняет экспериментально наблюдаемое увеличение скорости отслоения в зависимости от концентрации фосфата (58).
Двухступенчатая модель рабочего хода MVI предполагает, что при τχ1 плечо рычага разъединяется, и преобразователь перемещается назад в положение после хода. Таким образом, обратная нагрузка в MVI препятствует вращению преобразователя обратно в ориентацию PrePS, а не помогает ему, как в случае MV ( SI Приложение , рис. S16). Следовательно, предполагая, что в MVI только доинсультная конформация может связывать Pi, мы прогнозируем, что, в отличие от MV, скорость отделения MVI под нагрузкой не должна зависеть от концентрации фосфата в растворе.
Прогнозы.
Наше моделирование приводит к следующим предсказаниям: ( i ) Как предполагалось в другом месте (41), поворот плеча рычага MVI происходит в два этапа: сначала преобразователь достигает конформации R-состояния, а плечо рычага направлено в сторону зазубренный конец F-актина и отсоединен от двигателя; впоследствии несвязанное плечо рычага вращается, пока не достигнет конформации R-состояния. Первый шаг лишь слегка зависит от обратной нагрузки ( SI Приложение , рис.S6 – S8). Отсоединение плеча рычага от моторного домена после первого шага указывает на то, что, в отличие от MV (58), добавление Pi в раствор не должно приводить к более быстрому отрыву от F-actin оттянутого назад MVI. ( ii ) Петля, вокруг которой вращается плечо рычага, близка к N785. ( iii ) Энергия плеча рычага во время вращения свидетельствует о том, что ему мешают полностью растянуться назад. Кроме того, он по существу не связан с θ≈2,25 рад до θ≈π / 4 и только в последней части захватывается двигательной областью.( iv ) Поворот плеча рычага MVI происходит в основном на стороне актина. ( v ) Типичное время поворота из состояния U в состояние R для плеча рычага длиной 7,5 нм составляет около 0,2 мс. Это согласуется со схемой, в которой короткие шаги предпринимаются всякий раз, когда отстающая, свободная головка связывает F-actin до того, как завершается вращение несвязанного плеча рычага, то есть перед вторым этапом перехода PrePS → R.
Выводы
Мы изучили ход мощности MVI, смоделированный как переход из состояния PrePS в состояние R, сосредоточив внимание на динамике, связанной с преобразователем, ins2 и доменом IQ.
Наше моделирование позволяет сделать два основных вывода: ( i ) Рабочий ход MVI происходит в два этапа. На первом этапе двигательная область претерпевает переход PrePS → R, характеризующийся движением преобразователя, а затем плечо рычага поворачивается в конформацию R-состояния. ( ii ) Во время вращения плечо рычага в значительной степени отсоединяется от области двигателя. Вращение плеча рычага является стохастическим и происходит в основном на стороне F-актина.
Наши результаты основаны на CG-модели перехода PrePS → R, в которой ( i ) боковые цепи остатков не моделировались, ( ii ) структуры MVI с высвобождением Pi и АДФ были игнорируется, ( iii ) роль F-актина (в моделировании, проведенном при обратной нагрузке) аппроксимируется некоторыми ограничивающими потенциалами, ограничивающими внешние степени свободы MVI, и ( iv ) присутствие TH определяется моделируется как обратная нагрузка 6 пН, направленная параллельно F-актину.Хотя мы считаем, что некоторые ограничения могут быть ослаблены в будущих исследованиях, мы не ожидаем, что на наши выводы повлияет более полное описание силового удара MVI.
Материалы и методы
Приготовление состояний PrePS и R.
Мы подготовили состояния PrePS и R, объединив несколько структур PDB. Последовательность и две структуры на рис. 2 показывают, что модель MVI состоит из моторного домена, плеча рычага до остатка K834 домена IQ и двух связанных CaMs.
Структуры PrePS и R до остатка H786 (расположенного в середине ins2; рис. 2 A ) были взяты из структур PDB 4ANJ (41) и 2BKI (33) соответственно. Поскольку дистальные части структур PDB 4ANJ и 2BKI были либо частично замещены GFP (4ANJ), либо решены только частично (2BKI), мы завершили нашу модель, используя ins2, домен IQ и связанные CaM структуры PDB 3GN4 (23). . Чтобы создать непрерывную и бесшовную модель, остатки 776–816 (большая часть ins2 и начало домена IQ; рис.2) 4ANJ и 2BKI были наложены на одни и те же остатки в 3GN4, в результате чего среднеквадратичное значение составляло 0,38 Å и 0,47 Å соответственно. Результирующая структура PrePS (R) на рис. 2 B (рис. 2 C ) использовалась в нашем моделировании.
Эксперименты (8, 59) предполагают, что плечо рычага MVI претерпевает следующую последовательность переходов: ( i ) Из состояния PrePS, связанного с АДФ и фосфатом, в конформацию, в которой плечо рычага не перемещается, но моторный домен теперь подготовлен для высвобождения фосфата (состояние PiR) (59).( ii ) После высвобождения фосфата головка MVI достигает АДФ-связанного состояния. Во время этого перехода происходит большая часть качания, но конформация плеча рычага не соответствует таковой в состоянии R (8) (см. Ссылку 19 для крио-ЭМ изображения). ( iii ) После выпуска ADP двигатель наконец переходит в состояние R.
Мы устанавливаем PrePS в качестве начального состояния и R в качестве конечного состояния, игнорируя промежуточные продукты, которые либо плохо характеризуются структурой (связанный с ADP), либо близки к нашему начальному (PiR) или конечному (связанный с ADP) состояниям.
Мы не моделировали F-actin явно. Хотя привязка к дорожке влияет на то, как MVI проходит через цикл (5), мы полагаем, что это не оказывает существенного влияния на наши выводы, поскольку они зависят от конформационного перехода конвертера и только в меньшей степени от того, что запускает движение конвертер. По той же причине мы предполагаем, что отсутствие промежуточных состояний в нашей модели не окажет существенного влияния на наши выводы.
CG Модель.
Мы создали CG-представление MVI, в котором каждая аминокислота представлена как одна гранула с центром в положении Cα. Мы использовали самоорганизованную полимерную (СОП) версию CG-модели (60, 61), которая описана в SI Приложение , раздел 1. Здесь важно то, что характер взаимодействия между двумя гранулами зависит от их расстояния. в родном состоянии. Если расстояние между ними в нативном состоянии ниже RC = 8Å, и если они находятся на расстоянии не менее трех остатков друг от друга в последовательности, две бусины взаимодействуют посредством притягивающего потенциала типа Леннарда-Джонса с минимумом на их естественном расстоянии. и глубиной скважины ϵNAT = 2 ккал / моль.Связность цепи обеспечивается неограниченно растяжимым нелинейным упругим потенциалом между последовательными бусинами в последовательности. Взаимодействие между всеми остальными парами шариков чисто отталкивающее, с силой отталкивания при диаметре шариков σ = 3,8 Å, равной ϵNNAT = 1 ккал / моль.
PrePS → R переход.
Чтобы смоделировать переход PrePS → R, мы использовали метод, описанный в ссылке. 61 с небольшими вариациями, которые согласовывались с результатами полностью атомистического молекулярно-динамического моделирования MVI, выполненного с помощью специального компьютера Anton ( SI Приложение , рис.S1). Этот метод (подробности см. В приложении SI , раздел 1) использует энергетическую функцию, которая одновременно включает нативные взаимодействия в PrePS и в R-состояниях, и имитирует эффект высвобождения фосфата, ослабляя взаимодействия, связанные с PrePS-состоянием, таким образом облегчая переход PrePS → R.
Мы сгенерировали 96 траекторий за достаточно долгое время, чтобы гарантировать, что MVI завершил переход PrePS → R. В этих симуляциях мы не ограничивали моторную сферу.Следовательно, полное вращение MVI было удалено с использованием алгоритма Кабша (62, 63) для выравнивания моторного домена перед преобразователем (остатки 1–703 до конца спирали Sh2) с моторным доменом R-структуры, которые ранее были выровнены с актин-связанным миозином II из PDB 1MVW. После выполнения этой регулировки отслеживалось вращение плеча рычага. Область преобразователя и плечо рычага, которые подвергаются наибольшему изменению, не учитывались при выравнивании.
Мы также построили 96 траекторий с обратной нагрузкой 6 пН.В этом случае, чтобы избежать движения твердого тела MVI из-за внешней силы, мы имитируем эффект F-actin, ограничивая положение некоторых остатков моторного домена (см. SI Приложение , раздел 1 для деталей). Мы не выполняли выравнивание до отслеживания вращения плеча рычага. Мы обнаружили, что выполнение такого выравнивания не влияет на качество наших результатов ( SI Приложение , Рис. S17), указывая тем самым, что детали интерфейса актомиозина вряд ли будут влиять на качание несвязанного плеча рычага.Наконец, мы выполнили 32 моделирования состояния PrePS, не вызывая перехода. Эти модели обсуждаются в Приложении SI .
Благодарности
Мы признательны Йонатану Голдтзвику за критические беседы. Мы благодарны Мэтью Капориццо, Йельского университета Голдмана, Наото Хори, Синь Ли, Сумит Синха и Хуонг Ву за полезные комментарии. Благодарим анонимного рецензента за подробные комментарии. Компьютерное время Антона было предоставлено Питтсбургским суперкомпьютерным центром (PSC) через грант R01GM116961 от Национальных институтов здравоохранения.Аппарат Антона в PSC был щедро предоставлен D. E. Shaw Research. М.Л.М. благодарит команду PSC за помощь в настройке моделирования на Антоне. Авторы благодарят Техасский центр передовых вычислений при Техасском университете в Остине и суперкомпьютерные средства Университета Мэриленда за предоставление вычислительных ресурсов. Эта работа была поддержана Национальным научным фондом (NSF) в рамках грантов CHE 16-36424 и CHE 16-32756, а также Центром инженерной механобиологии NSF (15-48571).
Сноски
Автор: M.L.M. и D.T. разработали исследования; М.Л.М. проведенное исследование; М.Л.М. и D.T. проанализировали данные; и М.Л.М. и Д.Т. написали статью.
Авторы заявляют об отсутствии конфликта интересов.
Эта статья представляет собой прямое представление PNAS.
Эта статья содержит вспомогательную информацию на сайте www.pnas.org/lookup/suppl/doi:10.1073/pnas.1615708114/-/DCSupplemental.
локомоторных движений ног, вызванных ритмическими движениями рук у людей
Цитата: Sylos-Labini F, Ivanenko YP, MacLellan MJ, Cappellini G, Poppele RE, Lacquaniti F (2014) Локомоторные движения ног, вызванные Ритмическими движениями рук у людей.PLoS ONE 9 (3): e. https://doi.org/10.1371/journal.pone.00
Редактор: Роберт Дж. ван Бирс, Университет VU Амстердам, Нидерланды
Поступила: 4 декабря 2013 г .; Принята к печати: 4 февраля 2014 г .; Опубликован: 7 марта 2014 г.
Авторские права: © 2014 Sylos-Labini et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Эта работа была поддержана Министерством здравоохранения Италии, Министерством университетов и исследований Италии (проект PRIN), Итальянским космическим агентством (гранты DCMC, CRUSOE и COREA) и программой Европейского союза FP7-ICT (грант MINDWALKER № 247959 и грант AMARSi № 248311). Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Люди качают руками автоматическим стереотипным образом во время передвижения, сочетая движения верхних и нижних конечностей с целочисленным соотношением частот, как это делают животные во время четвероногих движений [1] — [4]. Эти движения рук характерны для ходьбы, бега, ползания, плавания, лазания и других походок, но они не являются обязательными (как при ходьбе со скрещенными руками). Движения рук во время локомоции могут быть результатом пассивной механической связи с движениями туловища и плеч, а также активных двигательных стратегий, направленных на снижение общих затрат энергии [1], [5], [6] или повышение стабильности походки [7], [8] ].Ритмическая активность мышц рук и плеч присутствует при вертикальной ходьбе [9] — [12], даже когда руки неподвижны во время ходьбы [9], [13], что указывает на влияние центральной двигательной программы. Координация между руками и ногами во время передвижения человека имеет много общего с таковой у четвероногих, включая опору на проприоспинальные связи [2], [14] — [19]. У животных координация между конечностями также может отражать надспинальный контроль; таким образом, нейроны задних конечностей в моторной коре головного мозга кошек реагируют на изменения движений передних конечностей во время локомоции [20].
Связь между конечностями у людей ранее изучалась путем вызова рефлексов в одной конечности и наблюдения за степенью, в которой движение другой конечности модулирует выражение рефлекса [21] — [24]. Недавно было показано, что активные движения рук увеличивают задействование мышц ног во время субмаксимального шага лежа [25]. Утверждалось, что лучшее понимание механизмов, лежащих в основе координации четырех конечностей, может иметь важные последствия для локомоторной реабилитации при некоторых нейромоторных расстройствах [22], [25] — [27].
У четвероногих, движения передних конечностей могут способствовать или даже запускать шаги задних конечностей, что согласуется с сочетанием шейных и пояснично-крестцовых центральных генераторов паттернов (CPG) [19], [28] — [31]. Однако менее ясно, может ли такое облегчение также проявляться у людей, которые предположительно имеют более слабую связь между движениями верхних и нижних конечностей в связи с эволюцией двуногого передвижения и необходимостью освобождать верхние конечности для выполнения манипулятивных задач. Здесь мы попытались выявить облегчение шага нижними конечностями движениями верхних конечностей, попросив участников ритмично двигать руками над головой, как при ходьбе руками.Мы предположили, что эти движения рук могут запускать автоматические чередующиеся движения ног, вызывающие ассоциации с локомоторными паттернами. Мы использовали протокол воздушного шага, поскольку было показано, что, по крайней мере, в случае тонической сенсорной стимуляции или спинномозговой электромагнитной стимуляции, автоматические движения ног легче вызвать при отсутствии нагрузки на конечности и контроля равновесия [32] — [ 35].
Методы
Участников
В первой серии экспериментов мы отобрали 33 здоровых добровольца, не имевших отношения к цели экспериментов (возрастной диапазон 23–50 лет, 18 мужчин и 15 женщин, длина ног 0.83 ± 0,04 м [среднее ± стандартное отклонение], рост 1,72 ± 0,10 м, вес 68 ± 7 кг) на наличие ответов ног. Девять из этих субъектов (возрастной диапазон 25–45 лет, 4 мужчины и 5 женщин, средняя длина ног 0,86 ± 0,05 м, рост 1,73 ± 0,10 м, вес 69 ± 10 кг), у которых заметные движения ног могли быть вызваны движениями рук. и те, кто смог вернуться в лабораторию для участия в дополнительных сеансах, были отобраны для подробных кинематических и ЭМГ записей во второй серии экспериментов. Исследования соответствовали Хельсинкской декларации, и мы получили информированное согласие всех участников в соответствии с процедурами Комитета по этике Фонда Санта-Лючия.
Опытная установка
Мы уменьшили действующие силы гравитации в плоскости движения нижних конечностей с помощью экзоскелета, расположенного горизонтально (патент Италии № Rm2007A000489). Система подробно описана в [35] — [37]. Вкратце, субъект лежал на правом боку, каждая нога была подвешена в независимом экзоскелете, что позволяло выполнять движения сегментов с низким коэффициентом трения и инерции (рис. 1А). Система нейтрализует составляющую силы тяжести, нормальную к поверхности лежа.Длина телескопического бедренного сегмента экзоскелета регулировалась в соответствии с длиной бедра испытуемого, а нога прикреплялась к экзоскелету с помощью манжеты таким образом, чтобы обеспечить наилучшее совмещение осей вращения. тазобедренных и коленных суставов с таковыми экзоскелета. Сегмент стопы оставался безудержным. Чтобы обеспечить удобную ширину ступеньки, мы также отрегулировали угол между двумя ногами, наклонив конструкцию, которая поддерживает верхнюю часть экзоскелета по отношению к кушетке.Верхняя часть тела испытуемого была зафиксирована посредством фиксации груди и плеч, а голова покоилась на подушке. Несмотря на то, что передне-задние движения туловища были ограничены, опора для бедра могла скользить по передне-задним направляющим кушетки, что позволяло вращать таз. Беговую дорожку (EN-Mill 3446.527, Bonte Zwolle BV, Нидерланды) наклонили на 90 ° и поместили на расстоянии вытянутой руки от плеч испытуемого перпендикулярно его / ее телу. Комфортное расстояние (достаточно большое, чтобы обеспечить достаточное сгибание / разгибание плеча и локтя, но не слишком большое, чтобы обеспечить легкий контакт с беговой дорожкой) от плеча до полотна беговой дорожки составляло около 80% от общей длины верхней конечности (предплечье + плечо). ) в любых условиях.В основной серии экспериментов полотно беговой дорожки перемещалось в сагиттальном (передне-заднем и задне-переднем) направлении относительно лежащего испытуемого (рис. 1А). В дополнительном эксперименте беговую дорожку вращали так, чтобы ее ремень двигался во фронтальном (срединно-латеральном) направлении относительно испытуемого.
Рисунок 1. Экспериментальная установка.
A — схематическое изображение горизонтальной системы поддержки веса тела. Испытуемый лежал на правом боку, каждая нога была подвешена в экзоскелете, позволяющем двигаться сегментами с низким коэффициентом трения.Оружие не было ограничено и не поддерживалось. Голова покоилась на подушке, закрепленной на горизонтальном ремне шириной 10 см (не показан) таким образом, чтобы нижняя часть руки двигалась беспрепятственно. B — пример передне-заднего смещения стопы (маркер MP), вызванного ходьбой (представлен смещением маркера DP). FW — вперед, BW — назад. L обозначает пешую экскурсию. Вертикальные пунктирные линии указывают начало движений рук и ног. Обратите внимание на разницу между периодами цикла стопы и руки (T 1 и T 2 , соответственно).C — круговая диаграмма, показывающая процент субъектов (n = 33), у которых ходьба вручную вызвала значительные (L> 10 см) смещения стопы, и субъектов с небольшим эффектом или без него (L <10 см).
https://doi.org/10.1371/journal.pone.00.g001
Протокол
Мы попросили участников дотянуться до беговой дорожки над головой и ритмично перемещать руки на движущемся ремне беговой дорожки, как если бы они ходили по рукам. Руки были неограниченными и без опоры (голова опиралась на небольшую подушку, закрепленную на горизонтальном ремне шириной 10 см таким образом, чтобы нижняя часть руки двигалась беспрепятственно).Чтобы «ходить» по беговой дорожке, участникам нужно было дотянуться до поверхности и активно вытянуть руку. Чтобы снизить уровень внимания, уделяемого двигательной задаче, испытуемых также просили выполнять в уме арифметические операции (отсчет вслух по семеркам) на протяжении каждого испытания [33]. Мы не давали никаких инструкций относительно позы, которую необходимо поддерживать телом и нижними конечностями. Изменяя скорость беговой дорожки в разных испытаниях, мы могли соответственно изменять скорость движений рук.Лента беговой дорожки могла смещаться в передне-заднем (а-р), задне-переднем (р-а) или срединно-боковом (m-l) направлениях в разных испытаниях. Протокол и инструкция были следующие. Сначала испытуемого просили вытянуть обе руки над головой, не касаясь беговой дорожки (исходное положение). Примерно через 2–3 секунды полотно беговой дорожки начало двигаться с заданной скоростью, и началась запись. Затем (примерно через 3–5 с) экспериментатор называл испытуемому случайное число, и он / она начинал считать и «ходить вручную» на беговой дорожке.Продолжительность каждого испытания составляла ∼1–2 минуты с минимум 2-минутным отдыхом между испытаниями. В конце испытания экспериментатор сказал испытуемому, чтобы он прекратил считать, и отключил движение полотна беговой дорожки. Прекращение ходьбы вручную не было зафиксировано в основном протоколе (таблица 1).
Все участники (n = 33) были первоначально проверены на наличие автоматических движений ног, вызванных ходьбой вручную со скоростью 1 и 2 км / ч в обоих направлениях: a-p и p-a. После завершения теста мы спросили испытуемых, заметили ли они появление движений ног во время ходьбы вручную.Эти сеансы записывались на видео, без автоматического захвата движения и сбора данных ЭМГ, средний ход стопы приблизительно оценивался с помощью линейки (в среднем 28 ± 24 см, диапазон 3–85 см).
Затем мы собрали подробные кинематические данные и данные ЭМГ в 2 дополнительных сеансах (таблица 1), проведенных в разные дни, у 9 испытуемых из тех, кто продемонстрировал заметные автоматические движения ног (ход стопы> 20 см) в ответ на воздействие рук. ходьба во время первоначального тестирования. На одном занятии участники ходили по беговой дорожке вручную с разной скоростью в направлении a-p (0.5, 1, 2, 3 и 4 км / ч в произвольном порядке), а в направлении р-а со скоростью 2 км / ч. В заключительной части этого занятия были проведены два дополнительных теста. В первом тесте мы просили испытуемых генерировать в воздухе шагающие движения рук в собственном темпе в п-направлении, а во втором тесте мы просили испытуемых выполнять движения ногами добровольно в течение 20–30 с (без движений рук. ) лежа в аппарате, как и раньше. Общая продолжительность этого экспериментального сеанса составляла ~ 2 часа.
На другом сеансе было проведено четыре дополнительных эксперимента на подгруппах субъектов (таблица 1).В одном эксперименте мы попросили испытуемых выполнить ручную ходьбу со скоростью 1 км / ч в срединно-латеральном (m-l) направлении. С этой целью беговая дорожка была размещена перпендикулярно первому этажу с движением ремня снизу вверх, что соответствовало ходьбе правой рукой. В другом эксперименте мы также записали движения ног после прекращения ходьбы вручную (три испытания со скоростью 1 км / ч в направлении AP для каждого испытуемого): участников проинструктировали прекратить ходьбу руками (сохраняя неподвижное положение руки), когда беговая дорожка была арестована, но счет продолжал.В третьем эксперименте мы проверили эффект временного блока шаговых движений ног, вызванного ходьбой вручную. С этой целью экспериментатор (находящийся позади испытуемого, невидимый для него / нее) блокировал на несколько секунд ноги испытуемого примерно в центральном положении их экскурсии во время ходьбы руками в п-направлении со скоростью 1, 2 и 3 км / ч. (произвольно выбранные скорости, 42 испытания по 6 испытуемым). Блок был получен путем ручного удерживания обоих сегментов голени экзоскелета. В четвертом эксперименте мы проверили потенциальные механические эффекты смещения бедра на движения ног.С этой целью экспериментатор вручную смещал нижнюю часть туловища испытуемого вперед и назад на ~ 5 см (сравнимо со средней амплитудой смещений туловища, измеренной во время ходьбы вручную в предыдущем сеансе, см. Результаты), в то время как испытуемый оставался пассивным.
Запись данных
Мы записали кинематические данные с двух сторон на частоте 100 Гц с помощью системы Vicon-612 (Оксфорд, Великобритания) с девятью камерами, расположенными по всей системе. Инфракрасные отражающие маркеры (диаметр 1,4 см) были прикреплены с каждой стороны пациента к коже над следующими ориентирами: конец 3-й дистальной фаланги (DP) кисти, запястье, локоть, плечево-плечевой сустав, большой вертел ( GT), латеральный надмыщелок бедра (LE), латеральная лодыжка, пятка и пятый плюснефаланговый сустав (MP).Маркер GT на правой стороне тела не удалось записать (поскольку объект лежал на этой стороне). 20-сантиметровые палочки с двумя маркерами были прикреплены к GT и LE левой ноги, и положения GT и LE были реконструированы как средняя точка между этими двумя маркерами.
ЭМГ-активность регистрировалась с двух сторон с помощью поверхностных электродов одновременно от 13 мышц: локтевого сгибателя запястья (FCU), локтевого разгибателя запястья (ECU), длинной головки трехглавой мышцы плеча (TRIC), длинной головки двуглавой мышцы плеча (BIC), передней дельтовидной мышцы. (DELTa), задняя дельтовидная мышца (DELTp), полусухожильная мышца (ST), двуглавая мышца бедра (BF), медиальная широкая мышца бедра (Vmed), прямая мышца бедра (RF), передняя большеберцовая мышца (TA), медиальная икроножная мышца (MG) и камбаловидная мышца (SOL).Данные ЭМГ были записаны с помощью беспроводной системы ЭМГ Delsys Trigno (Delsys Inc., Бостон, Массачусетс), ширина полосы 20–450 Гц, общее усиление 1000 и оцифровано с частотой 2000 Гц. Выборка кинематических данных и данных ЭМГ была синхронизирована.
Анализ данных
Цикл походки определялся независимо для движений рук и ног как временной интервал между двумя последовательными максимумами ap-смещения маркера DP для рук и маркера MP для ног (T 1 и T 2 , соответственно, на рис.1Б). Были вычислены амплитуды угловых изменений плеча, локтя, запястья, бедра, колена и голеностопного сустава [17] и усреднены по ~ 10 циклам в течение последних 15–30 с каждого испытания. Направление вызванного шага ногой в воздухе (вперед или назад) оценивалось путем вычисления траектории знаковой площади стопы (маркер MP) в течение цикла (нормированной на ход стопы) и усреднения по всем циклам в испытании: (1) где x и y — координаты MP-маркера (рис. 1B), Δx — ход ступни в i-м цикле, t (i) и t (i + 1) — это начала циклов i и i + 1 ног, соответственно, и N — количество циклов в испытании.По этому критерию движение считалось вперед, если область была положительной, и назад, если область была отрицательной.
Чтобы оценить процент субъектов в исходной выборке из 33 участников, у которых ходьба руками вызвала значительные смещения стопы, мы установили порог 10 см (L) на основе амплитуды (размаха) стопы (маркер MP ) смещения, вызванные пассивными движениями бедра (см. Влияние колебаний туловища на движение ног в Результатах). Поэтому мы рассматривали колебания нижних конечностей с L <10 см как возможные из-за механического воздействия движений рук на движения ног.
Мы определили начало движения ноги как последнюю критическую точку перед первым ритмическим циклом с L> 10 см, где производная хода стопы по времени (маркер MP) изменила свой знак (рис. 2). Начало движения руки определялось, когда ход конечной точки кисти превышал 2 стандартных отклонения от среднего значения, рассчитанного в период отдыха в начале каждого испытания, потому что всегда были небольшие колебания конечной точки руки из-за неподдерживаемого исходное положение руки (см. протокол).
Рисунок 2. Инициирование шага.
Примеры задержки вызванного шага в воздухе от начала движения руки (со скоростью, указанной в верхней части рисунка) у трех разных испытуемых. От верх до низ : передне-заднее (ап) смещение (дисп.) Левой руки, ЭМГ-активность 6 мышц левой руки, смещения левого бедра и стопы, углы левого тазобедренного и коленного суставов и ЭМГ активность 7 мышц левой ноги. Вертикальные пунктирные линии указывают начало движений рук и ног.FCU, локтевой сгибатель запястья; ЭКЮ, локтевой разгибатель запястья; TRIC — длинная головка трехглавой мышцы плеча; BIC, длинная головка двуглавой мышцы плеча; ДЕЛЬТА, передняя дельтовидная мышца; DELTp, задняя дельтовидная мышца; ST, полусухожильная; BF, двуглавая мышца бедра; Vmed, Wastus medialis; RF, rectus femoris; TA, tibialis anterior; MG gastrocnemius medialis; SOL soleus. Обратите внимание на вариабельность начала шага ног у испытуемых.
https://doi.org/10.1371/journal.pone.00.g002
Сигналы ЭМГ были численно выпрямлены, отфильтрованы нижними частотами с использованием фильтра Баттерворта с нулевой задержкой 4 th с отсечкой 10 Гц.Кинематические и ЭМГ-данные были интерполированы по времени по отдельным циклам походки, чтобы соответствовать нормализованной базе времени из 100 точек, и усреднены по (~ 10) циклам. Для усреднения по субъектам мы использовали как ненормализованные (в мкВ), так и нормализованные (до максимального значения) данные ЭМГ. Для нормализованного метода огибающие ЭМГ принимались равными нулю, если максимальное значение было меньше 3 мкВ (что мы считали уровнем шума). Мы также проанализировали кинематику конечностей и сигналы ЭМГ в частотной области с помощью быстрого преобразования Фурье (‘fft.m ’в Matlab). Мы вычислили частотную составляющую с наибольшей амплитудой (пиковую частоту), ее фазовый сдвиг относительно начала цикла отрезка (ноль соответствует косинусной функции с нулевым временным сдвигом) и учтенный процент дисперсии (r 2 ). для этого компонента.
Когда соотношение между продолжительностью цикла руки и ноги составляло ∼1 (в диапазоне 0,95 ÷ 1,05), ипсилатеральное фазовое отставание ( IPL ) между верхними и нижними конечностями определялось с использованием методов, описанных ранее [17].Вкратце, относительное время начала цикла левой нижней конечности выражалось в процентах от цикла походки, определяемого последовательными контактами левой руки: (2) где t 1 — интервал времени между началом цикла левой ноги и событиями касания левой рукой. и T — продолжительность ручного цикла. В соответствии с этим методом латеральные модели походки (ипсилатеральный контакт верхней / нижней конечностей в аналогичных случаях) определяются при значении 0%, а модели диагональной походки (контралатеральный контакт верхней / нижней конечности в аналогичных случаях) определяются при значении 50%. .Промежуточные значения (~ 25%) соответствуют отсутствию спаривания конечностей.
Чтобы оценить результативность мысленных арифметических операций, экспериментатор аннотировал все числа, произносимые в ходе испытаний на четырех испытуемых. Для каждого испытания мы вычисляли процент ошибок как количество ошибочных чисел, деленное на общее количество чисел и умноженное на 100, и скорость счета как общее количество чисел, деленное на продолжительность испытания (в минутах). .
Моделирование механического воздействия движения бедра на движения ног
Мы рассмотрели потенциальные механические эффекты колебаний туловища на движение ног, используя как биомеханическое моделирование, так и влияние внешних движений туловища, оцененных в дополнительных экспериментах (Таблица 1).В частности, на основе этих двух подходов мы определили a posteriori — нижний порог экскурсий стопы, используемый для расчета процента «реагирующих» субъектов из начальных скрининговых экспериментов (см. Результаты).
Биомеханическая модель использовалась для моделирования чисто механического воздействия колебаний туловища на движения ног. Нижняя конечность была смоделирована как мульти-маятник с тремя жесткими однородными сегментами (рис. 3A): бедро, голень и стопа, с массой м T , м S и м F , длина L T , L S и L F , и момент инерции I T , I S и I 95 F 907.Тазобедренные и коленные суставы моделировались как шарниры без трения с линейными амортизаторами, коэффициенты демпфирования составляли b H и b K соответственно [38]. Кроме того, чтобы учесть пассивное эластичное сцепление за счет одно- и двухсуставных мышц, для каждого сустава мы рассматривали пассивный упругий суставной момент ( M H и M K ) как функцию сустава нижней конечности. углы, следуя модели Ринера и Эдриха [39]. Мы считали голеностопный сустав фиксированным, потому что мы не наблюдали каких-либо значительных угловых движений в этом суставе во время соответствующих экспериментов (см. Результаты). Рисунок 3. Биомеханическая модель. A — схематическое изображение биомеханической модели, используемой для оценки механического воздействия периодических смещений бедра a-p на движение ноги. Б — примеры пассивных движений нижних конечностей, вызванных относительно большими (10 и 5 см) колебаниями бедра у двух испытуемых. Верхние кривые представляют собой пассивные смещения бедра, вручную индуцированные экспериментатором, а нижние кривые представляют собой результирующие движения ног (смещения стопы a-p и углы суставов) и ЭМГ мышц ног.Пунктирные линии представляют собой прогноз, сделанный с использованием биомеханической модели. Обратите внимание на отсутствие активности ЭМГ во время движений нижних конечностей, вызванных пассивным движением бедра. C — среднее (+ SD) горизонтальное отклонение стопы и сустава (от пика до пика), вызванное пассивным движением бедра и ходьбой вручную (среднее значение для всех скоростей беговой дорожки), а также предсказанное моделью. Обратите внимание на значительно меньшие смещения стопы, вызванные пассивным движением бедра или оцененные по модели (L <10 см), по сравнению с перемещениями при ходьбе вручную (~ 60 см).Звездочки обозначают достоверные различия (p <0,05) с тестами множественного сравнения Tukey HSD. https://doi.org/10.1371/journal.pone.00.g003 Координаты центра масс каждого сегмента: (3) где x GT — это ap-смещение маркера GT, h S , h T и h F — соответствующие положения центра масс в процентах от длины сегмента, а φ A — угол лодыжки. θ 1 и θ 2 — это углы возвышения (обобщенные координаты) в системе отсчета горизонтального экзоскелета. Инерционные параметры сегментов конечностей оценивали на основе скорректированных уравнений регрессии антропометрических данных [40]. В формулировке Лагранжа кинетическая энергия E системы равна: (4) потенциальная энергия U , функция диссипации Рэлея D (для учета вязкости) и обобщенные силы Q 1 и Q 2 составляют соответственно: (5) (6) (7) где и φ H и φ K — углы тазобедренного и коленного суставов соответственно. Движение нижней конечности было получено путем решения уравнений Лагранжа: (8) Для решения уравнения (8) был использован метод Рунге-Кутты четвертого порядка. 8 с начальными условиями, соответствующими начальному статическому положению ( θ 1 и θ 2 углов) с M H = 0; M K = 0; . Описательная статистика включала средние значения ± стандартное отклонение среднего. Для проверки нормальности распределения данных использовался тест Шапиро-Уилка.Повторные измерения (RM) ANOVA использовали для сравнения средних значений между различными условиями основного протокола и для оценки влияния различных скоростей ходьбы руками в направлении a-p. Апостериорные тесты и анализ множественных сравнений были выполнены с помощью теста Tukey HSD (Honestly Significant Difference). Уровень статистической значимости был установлен на уровне 0,05. Исходные данные доступны по запросу у авторов. Авторы предоставляют в свободный доступ любые материалы и информацию, описанные в этой статье, которые могут быть обоснованно запрошены другими в целях академических некоммерческих исследований.Пожалуйста, свяжитесь с [email protected] или [email protected]. Мы протестировали 33 участника, чтобы оценить процент «отзывчивых» субъектов. С этой целью участников, лежащих в разгрузочном экзоскелете, просили ритмично перемещать руки на беговой дорожке над головой в переднезаднем направлении. Чтобы снизить уровень внимания, уделяемого двигательной задаче, участников всегда просили выполнять в уме арифметические операции (отсчет вслух по семеркам) на протяжении всего испытания.Мы обнаружили, что этот тип ходьбы вручную вызывал автоматические движения ног с ходом стопы L> 10 см у 58% испытуемых (19/33, рис. 1С). У этих отзывчивых субъектов вызванные движения ног были ритмичными, чередующимися между левой и правой ногой, и сохранялись, пока субъект продолжал ходить вручную. У остальных испытуемых из нашей выборки ходьба руками вызвала небольшие (L <10 см) движения ног или их отсутствие. При опросе в конце эксперимента многие отзывчивые испытуемые, казалось, не осознавали, что они двигали ногами во время ходьбы вручную.Следует подчеркнуть, что из-за позы при ходьбе руками обзор нижних конечностей существенно затруднялся. В нескольких анекдотических случаях, когда экспериментатор направлял внимание испытуемого на наличие движений ног во время выполнения задания, испытуемый выглядел удивленным и заявлял, что ноги двигаются «сами по себе». Тем не менее, если его просили подавить автоматические движения ног, субъект, как правило, мог сделать это добровольно. Мы провели углубленные эксперименты с подробной записью кинематических и ЭМГ данных у 9 из испытуемых.У этих субъектов мы обнаружили, что движения ног систематически вызывались движениями рук в ходе повторных тестов без признаков адаптации, по крайней мере, в течение периодов наблюдения, которые не утомляли субъектов (1-2 мин). Начало движений ног всегда следовало за движением рук с задержкой по времени, которая сильно варьировалась в зависимости от испытуемых и условий (рис. 2). Хотя было некоторое начальное движение (возможно, из-за механической передачи, см. Ниже) сегментов ног примерно в начале ходьбы вручную в каждом случае реакции, задержка устойчивого колебательного движения ноги относительно начала движений руки варьировалась между 0.07 и 30 с у этих испытуемых (в среднем 4,05 ± 7,04 с). Самые длительные задержки имели место при самой низкой скорости ходьбы вручную (0,5 м / с), но в целом задержка систематически не зависела от скорости. Таким образом, у двух испытуемых, тестируемых на скорости 1 м / с на рис. 2, задержки были совершенно разными. В большинстве случаев (34/45 испытаний) задержки составляли менее 3 с. Представленные ниже результаты относятся к установившемуся состоянию вызванных реакций ног, обычно в последние 15–30 с каждого испытания при ходьбе руками в р-направлении, ходьбе руками со скоростью 2 км / ч в направлении р и в воздухе руками. шагая. Еще одна важная особенность локомоторных движений — направление шага. Мы проверили, демонстрируют ли движения ног обратный шаг с обратным движением рук. Анализ показал, что площадь траектории стопы была положительной как во время ап (1,5 ± 2,1 см, 2 / см, диапазон 0 ÷ 5,6 см, 2 / см), так и па (1,3 ± 2,7 см 2 / см, диапазон 0 ÷ 8,9 см 2 / см) направления ручной ходьбы со скоростью 2 км / ч. Таким образом, вызванные ритмические движения были преимущественно вперед независимо от направления ходьбы руками. На рис. 4 представлены общие кинематические параметры (вызванных) движений рук и ног. Угловые движения плечевого и локтевого суставов существенно не различались (F (2,16) = 0,227, p = 0,80 для плеча и F (2,16) = 2,45, p = 0,12 для локтя, RM-ANOVA) в разных руках. -Условия ходьбы (скорость 2 км / ч, па 2 км / ч и шаг в воздухе). Угловая амплитуда лучезапястного сустава была значительно больше (p = 0,035, Tukey HSD) при ходьбе вручную в направлении p-a, чем при ходьбе вручную в направлении a-p (рис.4Б, панели слева). Относительные угловые движения в тазобедренных и коленных суставах во время вызванных автоматических движений ног у разных испытуемых были разными (рис. 4А). В среднем они были на ~ 30 ° меньше в коленном суставе (рис. 4B, p <0,05, Tukey HSD) по сравнению с таковыми при произвольном шаге ногой по воздуху (рис. 4B, правые панели). Рисунок 4. Общие параметры походки. A — палочные диаграммы движений левой руки и ноги у трех разных испытуемых при ходьбе руками со скоростью 3 км / ч в направлении a-p (слева) и произвольной (vol.) воздушный шаг (справа). B — средние (+ SD, n = 9) угловые амплитуды суставов рук и ног при разных скоростях ходьбы руками в направлении AP (слева) и различных условиях основного протокола: ходьба руками со скоростью 2 км / ч в направлении Pa , воздушный шаг рукой (рука как) и произвольный шаг ногой в воздухе. C — продолжительность цикла и пешая экскурсия а-р. D — частотное соотношение движений рук и ног. На диаграммах разброса каждая точка показывает частоту движений рук или ног в различных условиях для каждого отдельного участника.Пунктирными линиями обозначены целочисленные отношения частот плеча: ноги (1∶1 и 2∶1). Столбцы на правой панели представляют собой средние (+ SD) отношения частот руки / ноги для всех условий. https://doi.org/10.1371/journal.pone.00.g004 Угловое движение в голеностопном суставе было довольно небольшим во время вызванных движений ног и сравнимо с тем, что было зарегистрировано во время произвольного шага в воздухе (p≥0,11, Tukey HSD). Горизонтальный ход стопы и длительность цикла вызванных движений ног были сопоставимы с произвольными шагами ногами по воздуху (рис.4C, p≥0,052, Tukey HSD). Продолжительность цикла колебаний рук и ног монотонно уменьшалась со скоростью (F (4,32) = 54,4, p <0,00001 для рук и F (4,32) = 8,30, p = 0,00027 для ног, RM-ANOVA, рис. 4С). Однако, в отличие от того, что обычно происходит при вертикальной ходьбе [5], [41], соотношение частот между движениями рук и ног отличалось от 1 в большинстве случаев при ходьбе вручную и в значительной степени зависело от скорости ходьбы вручную ( F (4,32) = 11,8, p <0,00001). Действительно, если сопоставить частоту движений рук с частотой движений ног (рис.4D), точки данных для ходьбы вручную во всех условиях попадают между линиями регрессии 1: 1 и 2: 1, что указывает на то, что частота движений ног, как правило, ниже, чем частота движений рук. Ходьба вручную обычно ассоциировалась с низким уровнем ЭМГ-активности мышц ног, что соответствовало ненагруженным условиям горизонтальной позы и ограниченным движениям в голеностопном суставе (рис. 2,5,6). Действительно, общая активность ЭМГ была низкой даже при произвольном шаге по воздуху (см. Рис.6,7), что также включало разгрузку и ограниченные движения голеностопного сустава (см. Также [42]). Более того, у разных субъектов наблюдалась вариабельность в моделях модуляции активности мышц ног, связанных с ходьбой вручную (сравните двух субъектов на рис. 5). ЭМГ, как правило, модулируется ритмично синфазно с движениями ног в мышцах подколенного сухожилия (ST и BF) чаще всего, а в других мышцах нижних конечностей более спорадически. Таким образом, субъект s6 на фиг. 5B продемонстрировал заметную модуляцию медиальной широкой мышцы бедра (Vmed) и прямой мышцы бедра (RF), в то время как субъект s2 на фиг.2 показана модуляция медиальной икроножной мышцы (MG). Также изменение активности ЭМГ со скоростью несколько зависело от субъектов. Например, активность мышц подколенного сухожилия увеличивалась со скоростью у субъекта s1 на фиг. 5A, в то время как она снижалась со скоростью у субъекта s6 на фиг. 5B (и активность Vmed и RF увеличивалась со скоростью у этого субъекта). Рис. 5. Модели ЭМГ у двух репрезентативных субъектов для разных скоростей ходьбы руками. От верх до низ : передне-заднее смещение левой руки, ЭМГ-активность 6 мышц левой руки, смещение а-р левой стопы и ЭМГ-активность 7 мышц левой ноги.Обратите внимание на нелинейные изменения в ЭМГ-активности со скоростью: у испытуемого 1 (A) наблюдалось увеличение активности мышц задней поверхности бедра, в то время как у испытуемой 2 (B) наблюдалось уменьшение активности подколенных сухожилий и увеличение активности четырехглавой мышцы. Отметим также отсутствие заметной активности дистальных мышц ног. https://doi.org/10.1371/journal.pone.00.g005 Рис. 6. Усредненные по ансамблю (по субъектам) кинематические и ЭМГ паттерны во время ходьбы вручную. От слева до справа : ходьба рукой со скоростью 2 км / ч в направлении a-p, p-a, шаг рукой в воздухе и произвольный шаг ногой в воздухе.От вверху до внизу : углы суставов (среднее ± стандартное отклонение, n = 9) (A), ненормализованная (B) и нормализованная (C) конверты ЭМГ (черные линии указывают среднее значение, а пунктирные линии показывают среднее значение + SD ). Образцы построены в зависимости от нормализованного цикла ног. Обратите внимание на одинаковое время активности мышц подколенного сухожилия (примерно в начале цикла ноги) во всех условиях. https://doi.org/10.1371/journal.pone.00.g006 Рисунок 7. Координация движений рук и ног. A — кинематика рук и ног и ЭМГ у одного репрезентативного участника, выполняющего ручную ходьбу со скоростью 2 км / ч и произвольное воздушное шагание. В левой части каждой панели показаны колебания левой верхней и нижней конечностей, сопровождаемые соответствующей ЭМГ-активностью некоторых репрезентативных мышц. Серые области разграничивают циклы отдельных конечностей, определенные отдельно для рук и ног как время между двумя последовательными максимумами смещения a-p соответствующей конечной точки. В правой части каждой панели отображаются спектры Фурье, полученные из сигналов слева от них.Пунктирные линии показывают положение первых пяти кратных основной частоты (f1), для которых отображается объясненный процент отклонения (PV f1 ). B — средние (± SD, n = 9) пиковые частоты ЭМГ и соответствующие кинематические паттерны руки (верхний график) и ноги (средний график) для ходьбы руками с различной скоростью и для произвольного шага ногами в воздухе (об.). Нижний график представляет фазу (относительно начала цикла ноги) первой гармоники для ЭМГ ноги и углов подъема бедра и голени (ноль соответствует функции косинуса с нулевым сдвигом по времени).В этот анализ были включены только ЭМГ с пиковой активностью более 2 мкВ и PV f1 > 20%. https://doi.org/10.1371/journal.pone.00.g007 На рисунке 6 сравниваются кинематические модели ног и модели ЭМГ, ансамбль, усредненный по всем испытуемым при ходьбе руками (ап и па при 2 км / ч), рука воздушный шаг и произвольный воздушный шаг. Примечательно, что модель модуляции мышц подколенного сухожилия при ходьбе руками была аналогична таковой при произвольном шаге ногами в воздухе, со всплесками активности в конце замаха и в начале стойки в обоих условиях (см.рис.6). Кроме того, кинематика ног при ходьбе руками напоминала кинематику при произвольном шаге в воздухе, оба условия предполагали гораздо большие изменения углов бедра и колена, чем угол лодыжки. Угловой ход суставов ног был низким при шаге в воздухе (как вызванном, так и произвольном), особенно в голеностопном суставе (рис. 6). Чтобы количественно охарактеризовать связь между движениями рук и ног, мы вычислили величину и фазу преобразования Фурье кинематических и ЭМГ переменных (рис.7). Что касается движений ног, мы проанализировали только активность мышц задней поверхности бедра (BF и ST), потому что это были мышцы, более последовательно модулируемые во время ходьбы руками и произвольного шага по воздуху среди записанных. Рисунок 7 иллюстрирует кинематику и ЭМГ у одного репрезентативного участника, выполняющего ходьбу руками в направлении a-p со скоростью 2 км / ч и произвольные шаги в воздухе. В частности, эти примеры показывают, что, в отличие от вертикальной ходьбы [2], частоты колебаний рук и ног во время ходьбы не были заблокированы между собой в соотношении 1: 1.Эти взаимосвязи между движениями рук и ног были подтверждены путем анализа ЭМГ-активности проксимальных мышц руки и ноги, даже несмотря на то, что процент отклонений, объясняемых основной гармоникой, обычно был меньше для профилей ЭМГ по сравнению с кинематикой как для ходьбы вручную, так и для произвольной ноги. воздушный шаг (рис. 7А). Пиковая частота модуляции ЭМГ тесно коррелировала с таковой для соответствующей конечности, руки для мышц руки и ноги для мышц ног (рис.7Б, верхняя и средняя панели). Фурье-анализ также подтвердил предыдущее качественное наблюдение (рис.6) о том, что фаза активности мышц BF и ST при ходьбе вручную (рис. 7B, нижняя панель) была примерно такой же, как и при вертикальной ходьбе (около конца махов и начала движения). позиции [43]). Когда соотношение между движениями рук и ног составляло ∼1 (в диапазоне 0,95 ÷ 1,05, всего 13 испытаний для всех условий), ипсилатеральное фазовое отставание (уравнение 2) между верхними и нижними конечностями было 34 ± 8% (диапазон 22 ÷ 47), демонстрируя, таким образом, более «диагональную» походку (IPL около 50%), хотя в четырех из 13 испытаний участники не демонстрировали отсечения конечностей (IPL = 22–28%).«Боковая» походка (IPL ~ 0%) никогда не наблюдалась. Согласно литературным источникам [44], [45], доказательство того, что задача выполняется автоматически и выполняется с минимальным вниманием, обеспечивается тем фактом, что второстепенная задача выполняется с минимальным вмешательством. Чтобы оценить степень вмешательства в задачи, у четырех испытуемых мы сравнили выполнение мысленной арифметики (количество ошибок и скорость счета) в периоды отдыха и во время ходьбы руками. Ни процент ошибок (5 ± 2% и 7 ± 4% при ходьбе и покое соответственно), ни скорость счета (18.8 ± 7,5 мин ( −1 и 15,2 ± 5,4 мин ( −1 соответственно)) достоверно различались в этих условиях (p = 0,18 и p = 0,05, парные t-критерии), хотя результативность ментальной арифметики была немного хуже. при ходьбе вручную. Эти результаты показали ограниченное взаимодействие между двумя задачами, таким образом подтверждая гипотезу об автоматичности движений ног, возникающих во время ходьбы руками [44], [45]. Также следует отметить, что ментальная арифметика сводит к минимуму внимание испытуемых к движениям ног. Другой вопрос: являются ли эффекты направленными или вызваны общим увеличением возбудимости спинномозговых локомоторных цепей из-за сокращений мышц руки. Например, известно, что маневр Джендрассика облегчает воздушную походку ногами [33]. В частности, мы проверили, могут ли автоматические движения ног быть вызваны движениями рук, выполняемыми в направлении, отличном от направления нормальной ходьбы. С этой целью мы попросили четырех испытуемых выполнить ручную ходьбу в переднезаднем или срединно-латеральном направлении с одинаковой скоростью (рис.8). Экскурсии рук в направлении движения полотна беговой дорожки существенно не различались между двумя условиями (28 ± 6 см против 29 ± 8 см соответственно, p = 0,39, парный t-критерий, хотя пример на рис. 8 показывает немного меньше ручные экскурсии по направлению мл). Однако вызванные движения ног в основном наблюдались во время ходьбы вручную (смещения стопы составляли 49 ± 27 см против 4 ± 3 см, p = 0,021, парный t-критерий). Кроме того, не было обнаружено никаких существенных различий в выполнении мысленной арифметической задачи, количественно выраженной процентом ошибок и скоростью счета между ходьбой руками в направлениях a-p и m-l (p> 0.32 для обоих параметров, парные t-тесты), что соответствует аналогичному уровню автоматизма для ходьбы вручную в двух направлениях. Рис. 8. Специфичность вызванных движений ног при ручной ходьбе. Пример движений ног при ходьбе руками в переднезаднем (a-p, A) и срединно-латеральном (m-l, B) направлениях у одного репрезентативного субъекта (s9). Формат аналогичен рис. 5. R — правый, L — левый. Отметьте минуту, если какие-либо движения ног во время ходьбы рукой в направлении m-l (B). https://doi.org/10.1371/journal.pone.00.g008 Для дальнейшего исследования природы вызванных движений ног мы вручную заблокировали движения ног у шести испытуемых. На рисунке 9 показаны примеры вызванных ответов у четырех испытуемых: пунктирными линиями обозначен период временной блокады ног (прерывание и возобновление смещений стопы). В большинстве исследований (27/42, 64%) мы наблюдали ЭМГ-активность мышц задней поверхности бедра (ST, BF) в ответ на этот маневр.Заметной активности ЭМГ в других мышцах ног мы не наблюдали. В 17 испытаниях у субъектов наблюдалась тоническая реакция, состоящая из постоянной активации во время блока (см. Субъект s8 на рис. 9A), в то время как в 10 исследованиях мы наблюдали ритмический ответ, состоящий из взрывных эпизодов с частотой около 0,6 Гц (субъект s2 на рис. 9А). Таблица 2 содержит количество испытаний с тоническими и фазовыми ответами у каждого субъекта. В остальных 15 исследованиях не было явной ритмической или тонической ЭМГ-активности мышц подколенного сухожилия.Все испытуемые возобновили автоматические движения ног после отпускания ног, продолжая ходить вручную (рис. 9А). Рис. 9. Эффект временной блокады ноги и прекращения ходьбы руками. A — Кинематика верхних и нижних конечностей и ЭМГ-активность у двух испытуемых во время временной блокады ног, выполняемой экспериментатором вручную. Пунктирными линиями обозначен период преходящей блокады ноги: прекращение и начало смещения стопы. Один субъект (s8) продемонстрировал тоническую реакцию в мышцах ног во время блока, в то время как другой (s2) показал фазовую реакцию.Все испытуемые восстановили непроизвольные шаговые движения в воздухе после расслабления ног. Б — продолжение движений ног после прекращения ходьбы руками. Обратите внимание на ЭМГ-активность мышц ST и BF, связанную с несколькими постциклами движений стопы. https://doi.org/10.1371/journal.pone.00.g009 Мы также проверили устойчивость движений ног и / или ЭМГ-активности после прекращения ходьбы вручную (Таблица 1, три испытания на скорости 1 км / ч в направлении a-p для каждого субъекта).Постоянные движения ног с соответствующей активностью мышц задней поверхности бедра наблюдались после прекращения движений рук (фиг. 9B). Задержка прекращения колебательных движений ног относительно окончания движений руки варьировала от 0,6 до 46 с. В большинстве случаев (9/12 испытаний) задержка превышала 3 с (в среднем 10,05 с). Потенциальные механические эффекты колебаний туловища на движение ног оценивались с использованием как биомеханического моделирования, так и эффекта внешних движений туловища.Результаты моделирования представлены на рис. 3. При усредненных горизонтальных (от пика до пика) смещениях стопы, предсказанных моделью с использованием фактических движений бедер во время ходьбы руками в качестве входных данных, было 8 ± 7 см (все скорости были объединены вместе. , Рис. 3C). Для непосредственной оценки влияния колебаний туловища на движения ног (таблица 1) экспериментатор перемещал туловище испытуемого вперед и назад, пытаясь имитировать колебания бедер, связанные с ходьбой руками. В результате среднее смещение бедер составило 5.45 ± 1,41 см по сравнению со средним смещением 3,53 ± 2,49 см, связанным с ходьбой вручную (рис. 3C, левая панель). Важно отметить, что мы обнаружили, что амплитуда (от пика до пика) смещений стопы (маркер MP), вызванных пассивными движениями бедра, была относительно небольшой (от 2 до 10 см, в среднем 7,30 ± 3,41 см). Более того, в отличие от движений ног, вызванных ходьбой вручную, мы не наблюдали какой-либо заметной ЭМГ-активности в мышцах ног с пассивными движениями бедра (рис. 3B). Мы также обнаружили, что амплитуда горизонтального смещения стопы, предсказываемая моделью, существенно не отличалась от экспериментально наблюдаемой при пассивных движениях бедра (рис.3B и C, p = 0,67, Tukey HSD). В целом, движения стопы, вызванные ходьбой вручную, были значительно больше, чем те, которые экспериментально измерены с пассивным движением бедра, и предсказанные биомеханической моделью (для обоих сравнений p = 0,00017 Tukey HSD). То же самое справедливо и для угловых экскурсий суставов (рис. 3C). Экскурсии стопы, вызванные пассивным смещением бедра, не превышали 10 см (рис. 3С). Поэтому мы использовали этот предел (L = 10 см) в качестве нижнего порога движений ног, связанных с ходьбой вручную, чтобы идентифицировать «отзывчивых» субъектов из пилотных экспериментов (рис.1С). Мы обнаружили, что ритмичное движение рук на беговой дорожке над головой, как при ходьбе руками, часто вызывает автоматические чередующиеся движения ног у значительной части испытуемых. Эти движения ног имели некоторое сходство с произвольными шагами в воздухе и вертикальными движениями. Таким образом, время ЭМГ-активности подколенного сухожилия относительно цикла походки было одинаковым во всех трех условиях. Более того, как и при обычной ходьбе, частота движений ног увеличивается с увеличением частоты движений рук во время ходьбы вручную.Однако соотношение частот руки / ноги имеет тенденцию становиться больше единицы с увеличением скорости беговой дорожки, в отличие от фиксированного отношения 1 при нормальной ходьбе. Связь между активностью шейных мотонейронов, лежащих в основе ходьбы руками, и активностью пояснично-крестцовых мотонейронов, лежащих в основе движений ног, была предположительно непрямой, отсроченной и асинхронной, по крайней мере, в наших экспериментальных условиях. Ниже мы обсуждаем результаты в контексте возможных функциональных связей между шейными и поясничными сетями, которые могут быть ответственны за наблюдения. Наша установка позволяла относительно неограниченное движение ног в эквипотенциальной плоскости силы тяжести, в то время как ходьба руками выполнялась с контролируемой скоростью. Несмотря на свои преимущества для исследования ритмогенеза, установка имеет ограничения. Во-первых, положение пациента лежа приводит к асимметричным вестибулярным и тактильным стимулам к телу и к некоторой активности мышц руки против силы тяжести, действующей перпендикулярно длинной оси руки. Однако маловероятно, что эти факторы, связанные с положением, оказали значительное влияние на общие характеристики шага.Действительно, ранее было показано, что ходьба в этой наклонной установке очень похожа на ходьбу с вертикальной опорой веса тела [35], [36]. Кроме того, мы не думаем, что движения ног представляют собой стратегию, используемую для облегчения «вертикального» досягаемости рук к беговой дорожке в позднем «качании» цикла рук, поскольку движения ног не наблюдались во время среднебоковой ходьбы руками ( Рис.8). Наконец, ритмичные движения ног были постоянными в разных условиях, включая разные положения рук и наличие или отсутствие сил «реакции земли» при ходьбе руками (шаг рукой в воздухе, рис.4,6). Во-вторых, положение руки над головой соответствует «ходьбе по деревьям» [46] или плаванию ориентации руки [2], а не взмаху руки во время передвижения по земле. Тем не менее, колебания плеча a-p при ходьбе вручную (∼7 см) были сопоставимы с колебаниями при обычной ходьбе (∼5–10 см [47]). Также часто считается, что двуногие люди произошли от четвероногих предшественников (наземных или древесных [26], [46]). Одним из отличительных аспектов ходьбы на четвероногих приматах является использование диагональных пар межконечностей [16], [48] — [50], которые мы также наблюдали в наших экспериментах, когда соотношение между частотами рук и ног составляло 1∶1. В-третьих, мы изучали влияние движений верхних конечностей на движения нижних конечностей, когда верхние и нижние конечности не выдерживали нагрузки. В этих условиях голеностопный сустав, как правило, не задействован (рис. 4В), вероятно, из-за отсутствия сил нагрузки, что согласуется с предыдущими исследованиями с использованием тонических сенсорных стимуляций (посредством вибрации мышц или электростимуляции нервов), чтобы вызвать движение ног по воздуху. движения [32], [33]. Органолептическая обратная связь вносит существенный вклад в активацию дистальных мышц во время передвижения [51], [52], а также схемы формирования шаблона в крестцовом мозге [53], возможно, может быть инактивирован, когда вход от опорной поверхности отсутствует.Продолжительность цикла вызванных колебаний ног (в среднем 2–4 с, рис. 4C) была больше, чем при вертикальной ходьбе (1,1–1,6 с, в зависимости от скорости [54]), что согласуется с влиянием силы тяжести на маятник. поведение конечностей и с идеей, что локомоторный контроллер использует преимущества и адаптируется к пассивной динамике многосуставной системы [5], [36]. Мы использовали воздушный шаг в качестве модели для исследования ритмогенеза у людей, поскольку его проявлению в значительной степени способствует отсутствие внешнего сопротивления, и он задействует неизменные сенсорные входы [33], [34].Более того, известно, что многие особенности координации четвероногих рук и ног сохраняются при выполнении различных локомоторных задач у людей [16], [55], включая реципрокный паттерн влияний между координацией движения и ходьбы [56] или координацией четвероногих конечностей во время избегание препятствий [57]. Наконец, нельзя исключить механическую передачу движений рук и связанного с ними скручивания туловища на ноги, учитывая низкое сопротивление экзоскелета. Однако, используя внешние движения туловища и биомеханическое моделирование (см. Влияние колебаний туловища на движение ног в Результатах), мы показали, что пассивные смещения бедер, примерно сопоставимые с теми, которые регистрируются при ходьбе вручную, определяют движения ног намного меньшие, чем те, которые связаны с ходьба руками и без заметной модуляции ЭМГ-активности в мышцах ног.Вместо этого такая модуляция ЭМГ присутствовала во время ходьбы вручную (рис. 2,5,6) и часто продолжалась даже тогда, когда движения ног были временно заблокированы экспериментатором (рис. 9A) или после прекращения движений руки (рис. 9B). ). Таким образом, основная часть свидетельств указывает на преимущественно активную (нервную), а не пассивную (механическую) природу движений ног, вызванных ходьбой вручную. Маловероятно, что движения ног были вызваны произвольно во время ходьбы руками: испытуемые всегда занимались ментальной арифметикой и, когда их спрашивали, они казались не замечающими движения ног (которые они не могли видеть из-за позы).В самом деле, общепринято, что автоматические движения выполняются без явного внимания к деталям движения [58]. Кроме того, ходьба руками мало влияла на выполнение мысленных арифметических операций (процент ошибок и скорость счета), что свидетельствует о минимальном вмешательстве или отсутствии вмешательства в двигательную задачу и, следовательно, на автоматичность [44], [45] движений ног. Хотя реакции ног были предположительно автоматическими, они не были стереотипными. Таким образом, задержка начала колебаний ног относительно колебаний руки была переменной (обычно около 1-2 с, но иногда и намного дольше), как и соотношение между частотой колебаний ног и частотой колебаний руки (рис.4,7). Так же, как произвольная воздушная ходьба, также ходьба руками обычно ассоциировалась с низким уровнем ЭМГ-активности мышц ног из-за ненагруженных условий. Мышцы подколенного сухожилия были мышцами ног, более последовательно активировавшимися ритмично, но у некоторых испытуемых также ритмично модулировались четырехглавые мышцы и икроножные мышцы (рис. 2,5,6). Это можно объяснить важным вкладом рефлексов растяжения в мышцу подколенного сухожилия (особенно в конце маха) в контексте «пассивного» вклада [59].Более того, ограниченная активация более дистальных мышц может быть интерпретирована с точки зрения их более сильной зависимости от сенсорной обратной связи, связанной с нагрузкой на конечности, по сравнению с проксимальными мышцами (в контексте «активного» вклада центральных источников) [51], [52]. Когда экспериментатор временно блокировал движения ног (Таблица 2) или после прекращения движений рук (Рис.9), в разных испытаниях мы могли наблюдать один из 3 различных ответов на ЭМГ-активность ранее активных мышц: 1 ) ритмическая активность, 2) тоническая активность, или 3) не обнаруживаемая активность.Постоянство ритмической активности ног (рис. 9A, B) дополнительно поддерживает активную (нервную), а не пассивную (механическую) природу движений ног, вызванных ходьбой рук, и, таким образом, указывает на активацию схем генерации паттернов. Напротив, наличие тонической активности может зависеть от взаимосвязи между мышечным тонусом и движением. Примеров такой взаимосвязи много. Например, мышечная активность может быть продлена, когда движение конечности прерывается в фазе замаха или стойки, а чередующиеся всплески могут быть заменены тонической активностью, когда конечность остается неподвижной [60] — [62].Более того, эпидуральная стимуляция на уровне L5 позвоночника у децеребрированных кошек первоначально вызывает тоническую активность в мышцах задних конечностей, которая меняется на двигательную активность после 5-7 секунд стимуляции [34]. Кроме того, инициирование локомоции, вызванной стволом мозга, обычно сопровождается повышением постурального мышечного тонуса [63]. Наконец, разные формы передвижения человека могут быть связаны со специфическим мышечным тонусом [64]. О больших межиндивидуальных различиях у людей (Fig. 1C, 2,9) в задержке и реакции схемы генерации паттерна позвоночника на его активацию также сообщалось в предыдущих исследованиях [32] — [34]. Каким бы ни был точный механизм наблюдаемого явления, эти переменные особенности предполагают, что сигналы, относящиеся к движениям рук, не передают напрямую двигательные команды мышцам ног, а вызывают реакции, которые зависят от сенсорной обратной связи и состояния пояснично-крестцовой локомоторной схемы [65]. . Один из возможных путей для этих триггерных сигналов — через внутренние спинномозговые пути, связывающие шейные и пояснично-крестцовые нейроны. Наиболее известными из таких связей являются длинные нисходящие проприоспинальные нейроны, которые были продемонстрированы у людей [14].Однако, учитывая латентность ответов ног относительно колебаний руки, нельзя исключить супраспинальный вклад. Как нисходящие, так и восходящие связи между шейными и пояснично-крестцовыми CPG описаны у четвероногих млекопитающих. У этих животных связь между конечностями намного сильнее, чем у людей, но функциональное состояние этих связей зависит от задачи и контекста [18]. Наши результаты показывают, что, как и у четвероногих животных [19], [29] — [31], ритмичные движения верхних конечностей могут инициировать шаги нижних конечностей у двуногих людей.Более того, как у кошек (см. Рис. 2 в [31]), так и у людей (рис. 4,7), шагание задними конечностями с помощью передних конечностей часто характеризуется нецелым соотношением между движениями передних и задних конечностей. Более того, стоит подчеркнуть, что наблюдаемое явление конкретно связано с циклическими движениями рук. Например, сильное изометрическое сокращение мышц руки (как в маневре Jendrassik) может увеличить возбудимость рефлекторных путей [66], [67], но не вызывает колебаний ног [33].Кроме того, среднебоковые ритмические движения рук неэффективны для запуска шага ногами по воздуху (рис.8), что подтверждает гипотезу о том, что механизмы, лежащие в основе этого явления, связаны с движениями рук, специфичными для направления, а не с общим увеличением возбудимости двигательная схема ног из-за сокращений мышц рук. Действительно, раскачивание руки обычно происходит в направлении a-p во время нормального передвижения. Мэтт Брэди — Чикаго Связь и Эффективность — горячие слова, когда речь идет о качелях для гольфа.Если вы когда-нибудь брали уроки гольфа или просматривали видео на YouTube, скорее всего, вы слышали одно, если не оба упомянутых ранее. Давайте определим, что они означают в самых общих чертах, а затем погрузимся в то, как они применяются к игре в гольф. Связь определяется как отношения, в которых человек, вещь или идея связаны или связаны с чем-то другим, легко понять, правда? В игре в гольф на это чаще всего ссылаются на взаимоотношения между руками игроков и их телом. Это означает, что для большинства игроков мы хотели бы, чтобы руки устанавливали и поддерживали соединение на протяжении всего замаха, начиная со статической установки и заканчивая замахом назад и через точку контакта с мячом для гольфа.Из этого правила есть небольшие исключения, но их не так много, как вы думаете. На ум приходит Джастин Томас, у него очень высокие руки на вершине взмаха, и похоже, что его руки отрываются от его тела, и они это делают совсем немного. Но один из его первых ходов в даунстрим — снова соединить руки с телом и поддерживать эту связь через зону удара. Итак, сам собой напрашивается вопрос, как улучшить связь через качели? Есть много способов и вещей, на которых вы можете сосредоточиться, будь то правый локоть, заправленный в грудную клетку по адресу и удерживающий его близко к вашему телу, спине и во время замаха, или ощущение левой руки на груди в верхней части качели.Ответ таков: все, что помогает вам добиться желаемого повторяемого полета мяча, является правильным способом. Вторая часть этого поста посвящена эффективности, которая определяется как достижение максимальной производительности с минимальными затратами усилий или предотвращение расточительного использования определенного ресурса. Это возвращает нас к первой части сообщения. Связь и эффективность, на мои деньги, очень похожи, когда дело касается самих качелей, и у вас не может быть одного без другого. Да, они могут относиться к различным частям игры в гольф, но начните с упомянутого выше соединительного элемента, и я надеюсь, что это поможет прояснить любые нечеткие мысли, которые у вас были по этому поводу. Статистика
Доступность данных
Результаты
Пилотные эксперименты
Количественная оценка движений ног при ходьбе руками
Паттерны ЭМГ при ходьбе руками
Координация движений рук и ног
Оценка автоматичности
Эффект ходьбы вручную, зависящий от направления
Эффекты блокировки движений ног
Последствия прекращения ходьбы вручную
Влияние колебаний туловища на движение ног
Обсуждение
Методологические соображения
Предполагаемые механизмы
и эффективность в Golf Swing